【欧州の歴史】ヨーロッパ人がなんで容姿淡麗か
1 :世界@名無史さん:
なんかで見たんだけど、ヨーロッパ人が容姿淡麗なのは500〜1000年ぐらい前に意図的にブサイクを大量虐殺して、容姿淡麗な民族を作ろうとしたからみたいなんだけど、何事件か教えて下さい。
|
|
|
逆。ヨーロッパ人が世界の文化や文明を最初に制覇しちゃったから、
ヨーロッパ人の価値観が美の基準になっただけ。
美醜は単なる文化。
その証拠に、浮世絵の美人絵って当時の美人の基準だ。平安絵巻みたいな
うりざね型は今ではブスの代名詞だろ。目も細いし。
ヨーロッパ人の価値観が美の基準になっただけ。
美醜は単なる文化。
その証拠に、浮世絵の美人絵って当時の美人の基準だ。平安絵巻みたいな
うりざね型は今ではブスの代名詞だろ。目も細いし。
× 淡麗
○ 端麗
○ 端麗
欧米人に美人が多いのは歯並びを矯正するから。
よほど貧乏で無い限り、男も女も歯の矯正はする。
二重瞼で鼻が高いから、歯さえ直せば美男美女。(太らなければ)
逆に治さないと白人でもブ男やブスになるリスクを被るうえに、
自分は育ちが悪いです、と言っているようなもんだ。
日本人も100%とは言わないまでも、
歯並びさえ治せばたいていの人は美男美女になるんじゃないかな。
金待ちの子女はみんなそこそこいくだろ。
よほど貧乏で無い限り、男も女も歯の矯正はする。
二重瞼で鼻が高いから、歯さえ直せば美男美女。(太らなければ)
逆に治さないと白人でもブ男やブスになるリスクを被るうえに、
自分は育ちが悪いです、と言っているようなもんだ。
日本人も100%とは言わないまでも、
歯並びさえ治せばたいていの人は美男美女になるんじゃないかな。
金待ちの子女はみんなそこそこいくだろ。
縄文人は歯が小さくて歯並び良かったみたいだよ。
硬いモノを食べないからアゴ細くなり、歯並びが悪くなるんだよ。
アゴが細い方が最近では美しいって要素になっているこの矛盾。
アゴが細い方が最近では美しいって要素になっているこの矛盾。
日本人は歯の小さい縄文人と歯の大きい弥生人が混血したから歯並びが悪くなった
日本人の八重歯の発生率は異常に高い
逆に朝鮮人はエラが発達してるから八重歯があまり見られない
日本人の八重歯の発生率は異常に高い
逆に朝鮮人はエラが発達してるから八重歯があまり見られない
日本人が綺麗になったのは、進駐軍がレイプしてくれたおかげw
9 :世界@名無史さん:2012/08/22(水) 14:03:03.40 0
室伏アニキがうらやましい。
10 :世界@名無史さん:2012/08/22(水) 23:13:08.77 0
コーカソイドの先祖はエデンの園( ペルシャ湾、及び緑だったその周辺 ) で豊かで暖かい
環境で余裕があり、美人がモテるという性淘汰が発生した。
一方、ヨーロッパに4万年前に入った第一陣の人類は黒人状態だったが氷河期が終わって
ペルシャ湾も水没したのでヨーロッパをはじめコーカソイドたちは中央アジアや
北アフリカなどに広まった。
一方、モンゴロイドの先祖は極地に入り全身毛皮で容姿もクソも無く生きるのに精一杯で
性淘汰は起きなかった。こういうシナリオで学会で通らないか?
環境で余裕があり、美人がモテるという性淘汰が発生した。
一方、ヨーロッパに4万年前に入った第一陣の人類は黒人状態だったが氷河期が終わって
ペルシャ湾も水没したのでヨーロッパをはじめコーカソイドたちは中央アジアや
北アフリカなどに広まった。
一方、モンゴロイドの先祖は極地に入り全身毛皮で容姿もクソも無く生きるのに精一杯で
性淘汰は起きなかった。こういうシナリオで学会で通らないか?
12 :世界@名無史さん:2012/08/22(水) 23:47:01.39 0
もう、それらしい説は発表されてるわ
http://ja.wikipedia.org/wiki/%E9%87%91%E9%AB%AA
http://ja.wikipedia.org/wiki/%E9%87%91%E9%AB%AA
日本に来たとき天狗とか思われてた
中国でもこんな珍妙な生き物が居たのかと書いてなかったっけ
中国でもこんな珍妙な生き物が居たのかと書いてなかったっけ
中国には、トルコ系の人が沢山出入りしていたから、碧眼とか珍しくなかったのでは?
ウイグル系もあるな。金髪はあまーりいないだろうけど。
ウイグル系もあるな。金髪はあまーりいないだろうけど。
新疆ウイグル自治区内のタジク族
http://img.ifeng.com/res/200809/0904_446974.jpg
http://img.club.pchome.net/upload/club/other/2008/11/23/pics_ask4help_1227430148.jpg
http://www.fyjs.cn/bbs/attachments/Mon_0904/149_101254_d3e4112e6ffd939.jpg
http://www.fyjs.cn/bbs/attachments/Mon_0904/149_101254_2d556ff0557d05b.jpg
http://www.fyjs.cn/bbs/attachments/Mon_0904/149_101254_b16b1482421e972.jpg
http://www.fyjs.cn/bbs/attachments/Mon_0904/149_101254_85d7d445ac57b0c.jpg
http://www.fyjs.cn/bbs/attachments/Mon_0904/149_101254_d9c507de357f0e7.jpg
http://hiphotos.baidu.com/xiaopiggykk/pic/item/b75f1d19985ea5964aedbceb.jpeg
http://hiphotos.baidu.com/wudidehankesi/mpic/item/f15093fb0accf43ca8d311b4.jpg
http://hiphotos.baidu.com/wudidehankesi/mpic/item/6f4b1939c7151de0d56225aa.jpg
http://hiphotos.baidu.com/wudidehankesi/mpic/item/f3bb3c8351199e9d6c8119ae.jpg
http://img.ifeng.com/res/200809/0904_446974.jpg
{kind=link}
http://img.club.pchome.net/upload/club/other/2008/11/23/pics_ask4help_1227430148.jpg
{kind=link}
http://www.fyjs.cn/bbs/attachments/Mon_0904/149_101254_d3e4112e6ffd939.jpg
{kind=link}
http://www.fyjs.cn/bbs/attachments/Mon_0904/149_101254_2d556ff0557d05b.jpg
{kind=link}
http://www.fyjs.cn/bbs/attachments/Mon_0904/149_101254_b16b1482421e972.jpg
{kind=link}
http://www.fyjs.cn/bbs/attachments/Mon_0904/149_101254_85d7d445ac57b0c.jpg
{kind=link}
http://www.fyjs.cn/bbs/attachments/Mon_0904/149_101254_d9c507de357f0e7.jpg
{kind=link}
http://hiphotos.baidu.com/xiaopiggykk/pic/item/b75f1d19985ea5964aedbceb.jpeg
{kind=link}
http://hiphotos.baidu.com/wudidehankesi/mpic/item/f15093fb0accf43ca8d311b4.jpg
{kind=link}
http://hiphotos.baidu.com/wudidehankesi/mpic/item/6f4b1939c7151de0d56225aa.jpg
{kind=link}
http://hiphotos.baidu.com/wudidehankesi/mpic/item/f3bb3c8351199e9d6c8119ae.jpg
{kind=link}
トルコの王宮には白人の宦官がいたし、
千一夜物語には「フランク人」の嫁さんを迎えてウハウハ(死語)などという物語がかなりある。
文化が発達する以前から、白人の容姿は美しいと見なされていたのでは?
千一夜物語には「フランク人」の嫁さんを迎えてウハウハ(死語)などという物語がかなりある。
文化が発達する以前から、白人の容姿は美しいと見なされていたのでは?
キエフを滅ぼしたバトゥは
ロシア人の美しさに目を奪われた。
これまで会ってきたどの人種よりも美しかった。
そのロシア人の中でも特に見栄えのいいものを
眺めることがバトゥの楽しみになった。
つまり視姦が趣味になったのだ。
ロシア人の美しさに目を奪われた。
これまで会ってきたどの人種よりも美しかった。
そのロシア人の中でも特に見栄えのいいものを
眺めることがバトゥの楽しみになった。
つまり視姦が趣味になったのだ。
18 :世界@名無史さん:2012/08/23(木) 02:05:13.41 0
19 :世界@名無史さん:2012/08/23(木) 03:53:11.23 0
>>11
違う。
それは人種差別や偏見を含むためにその論に誰も反対しないだけ。
どの文化圏のどの人種が見ても引き締まった体躯の汗血馬はロバより美しい。
ライオンよりハイエナの方が美しいと思う文化は無い。
かつては西洋でも一般ではぽっちゃり型の方が好まれていた。
やせていれば出差に危険が伴うし、授乳・免疫などの問題で子供が育たない可能性がある。
何より、親の経済的脆弱性が問題とされる。
貴族はその点、経済的・育児的問題が無いために「やせている」事を競うようになった。
しかしこれは、ようするに女性が衣食住事足りた場合、
やはり現在の美の基準が最も正しい事を示す。
生命の維持と繁殖に問題が無いのならば、手足がすらりと長く
豊かな髪、細い腰に美を感じるのは普遍的な人間の本能。
西洋よりも遥かに古い文化圏の中近東、あるいは古代中国でもその価値観は変わらない。
違う。
それは人種差別や偏見を含むためにその論に誰も反対しないだけ。
どの文化圏のどの人種が見ても引き締まった体躯の汗血馬はロバより美しい。
ライオンよりハイエナの方が美しいと思う文化は無い。
かつては西洋でも一般ではぽっちゃり型の方が好まれていた。
やせていれば出差に危険が伴うし、授乳・免疫などの問題で子供が育たない可能性がある。
何より、親の経済的脆弱性が問題とされる。
貴族はその点、経済的・育児的問題が無いために「やせている」事を競うようになった。
しかしこれは、ようするに女性が衣食住事足りた場合、
やはり現在の美の基準が最も正しい事を示す。
生命の維持と繁殖に問題が無いのならば、手足がすらりと長く
豊かな髪、細い腰に美を感じるのは普遍的な人間の本能。
西洋よりも遥かに古い文化圏の中近東、あるいは古代中国でもその価値観は変わらない。
>>17
肝心のそのロシア人たちは、支配者のモンゴル人の形質である背が低い人を美しいとしたけど?
>>19
はあ?ぽっちゃり体型の女性を多くを描いた人もいるが?
現代の美の基準は、細すぎる体型を美としすぎ。
標準体型の+10%が統計的に、一番長生きとされているんだろ?
芸能人の体型で行くと、山田花子あたりが一番長生きの体型に該当する。
現在は、美の基準が不健康過ぎる。これじゃ、中国のてん足と変わらない。
肝心のそのロシア人たちは、支配者のモンゴル人の形質である背が低い人を美しいとしたけど?
>>19
はあ?ぽっちゃり体型の女性を多くを描いた人もいるが?
現代の美の基準は、細すぎる体型を美としすぎ。
標準体型の+10%が統計的に、一番長生きとされているんだろ?
芸能人の体型で行くと、山田花子あたりが一番長生きの体型に該当する。
現在は、美の基準が不健康過ぎる。これじゃ、中国のてん足と変わらない。
世界中を回って比較したわけではないが、テレビなどからの印象を加味すると、
モロッコからアフガニスタンあたりにかけての地域が最も容姿が優れていますね。
ヨーロッパ系は南欧と東欧はいいけど、中欧から北欧は駄目ですね。
モロッコからアフガニスタンあたりにかけての地域が最も容姿が優れていますね。
ヨーロッパ系は南欧と東欧はいいけど、中欧から北欧は駄目ですね。
女は色白がもてるのはどこも共通する文化であり
いわば文明だな
いわば文明だな
24 :世界@名無史さん:2012/08/24(金) 01:00:40.39 0
スレ主だけど、最近テレビで見た事件なんだよね。やっぱ情報薄くてわからないかぁ。ggってもそれらしいの出てこないし
>キエフを滅ぼしたバトゥは
>ロシア人の美しさに目を奪われた。
>これまで会ってきたどの人種よりも美しかった。
>そのロシア人の中でも特に見栄えのいいものを
>眺めることがバトゥの楽しみになった。
中世ロシア史とモンゴル帝国史関連の本だいぶ読んだんですけど聞いたことありません。
出典は?
>ロシア人の美しさに目を奪われた。
>これまで会ってきたどの人種よりも美しかった。
>そのロシア人の中でも特に見栄えのいいものを
>眺めることがバトゥの楽しみになった。
中世ロシア史とモンゴル帝国史関連の本だいぶ読んだんですけど聞いたことありません。
出典は?
27 :世界@名無史さん:2012/08/24(金) 01:05:49.97 0
中世ロシア史なんてどこで学べるんだ
>>27
無理矢理無視するなよw
無理矢理無視するなよw
逆に肌白を醜いとする文化を知っているだろうか
いやそれ以前にガングロを美しいと感じた人間を知っているだろうか・・・
いやそれ以前にガングロを美しいと感じた人間を知っているだろうか・・・
31 :<ヽ`∀´> ◆NIDA/olocc :2012/08/24(金) 02:30:15.06 0
あまり白いのはイヤだな。小麦色がいい。それと弥生系、朝鮮系はイヤだ。
縄文系の目がパッチリで多少ぽっちゃりで○ぽい(・ω・)で、ボインがいいな。
縄文系の目がパッチリで多少ぽっちゃりで○ぽい(・ω・)で、ボインがいいな。
33 :名無し募集中。。。:2012/08/24(金) 06:51:00.02 0
美人が多いのはロシアでも南よりのウクライナとかでしょ
34 :世界@名無史さん:2012/08/24(金) 10:04:47.02 0
リトアニアとかベラルーシだろ
35 :世界@名無史さん:2012/08/24(金) 10:15:56.78 0
いずれもアーリア人の血が濃い地区だな
37 :世界@名無史さん:2012/08/24(金) 12:12:01.36 P
ウクライナはロシアの西では?
ウチにいるロシア人社員たちによると、
少しラテン系が入ったのが好みのようだ。
肌の色は少し褐色が入ったくらい。
眼孔が深すぎず、目は大き過ぎず小さすぎず、目尻は切れ長。
唇が厚い。中肉中背で背が高い。
要するに、機内誌の化粧品の広告とかでよく見るちょっとコワイ顔のお姉さんだね。
あと健康度は重要で、どんな美人顔でも、
肌がきれいで歯並びが良くないとアウト。
東洋人の顔は美醜のモノサシの外のようで健康度が重要な判断基準になるらしい。
別に東洋人だから醜いとは思っていないようだ。
ウチにいるロシア人社員たちによると、
少しラテン系が入ったのが好みのようだ。
肌の色は少し褐色が入ったくらい。
眼孔が深すぎず、目は大き過ぎず小さすぎず、目尻は切れ長。
唇が厚い。中肉中背で背が高い。
要するに、機内誌の化粧品の広告とかでよく見るちょっとコワイ顔のお姉さんだね。
あと健康度は重要で、どんな美人顔でも、
肌がきれいで歯並びが良くないとアウト。
東洋人の顔は美醜のモノサシの外のようで健康度が重要な判断基準になるらしい。
別に東洋人だから醜いとは思っていないようだ。
38 :世界@名無史さん:2012/08/24(金) 12:20:16.55 0
ニーナ・ドブレフとか。
>>23
図書館行って「ロシア史」「中世ロシア」「ルーシ」あたりで検索すれ
図書館行って「ロシア史」「中世ロシア」「ルーシ」あたりで検索すれ
>>37
日本の80年代アイドルの歯並びは悪かったなw
そうやってわざと歯科矯正しないで、普段着のアイドルってのが当時の売りだったような気がする。
シベリア抑留者の記録を読んでいると、男でも特定の人がやはりモテるんだよな。
俺は文章による印象だと「やさ男」系統がモテている印象を持っているが、写真を見ていないから本当のトコは分からないw
日本の80年代アイドルの歯並びは悪かったなw
そうやってわざと歯科矯正しないで、普段着のアイドルってのが当時の売りだったような気がする。
シベリア抑留者の記録を読んでいると、男でも特定の人がやはりモテるんだよな。
俺は文章による印象だと「やさ男」系統がモテている印象を持っているが、写真を見ていないから本当のトコは分からないw
41 :世界@名無史さん:2012/08/24(金) 14:42:11.54 P
>>38
こんな顔の子が事務所におったとき、確かに男どもの目線を集めとったわ。
この辺(ロシア西部)の女の子は、
俺らの目から見ると美人がデフォルトでブスがほぼ皆無。
当のロシア男にしてみると、美人に越したことは無いが、
それは大した問題では無く、健康度と性格が重要らしい。
ウチのロシア人たちの彼女の写真を見ると、
確かに中肉中背、お肌つるつる、歯並びバッチリだ。
(恥ずかしげもなく、彼女とのツーショットを壁紙にしている。
まあ、結局どれもモデルか女優かアイドル並なのだが。)
こんな顔の子が事務所におったとき、確かに男どもの目線を集めとったわ。
この辺(ロシア西部)の女の子は、
俺らの目から見ると美人がデフォルトでブスがほぼ皆無。
当のロシア男にしてみると、美人に越したことは無いが、
それは大した問題では無く、健康度と性格が重要らしい。
ウチのロシア人たちの彼女の写真を見ると、
確かに中肉中背、お肌つるつる、歯並びバッチリだ。
(恥ずかしげもなく、彼女とのツーショットを壁紙にしている。
まあ、結局どれもモデルか女優かアイドル並なのだが。)
外国でも少数にしても優男志向はあると思うが、優男って日本以外だとなんて呼ばれてるの?
英語ですら思いつかないわ
英語ですら思いつかないわ
アメリカでは、明らかに筋肉むちむちの方に志向が向いているな。
なんで日本には優男志向が強いんだろう?
なんで日本には優男志向が強いんだろう?
トーレスだって筋肉ムキムキじゃなくても普通の人に比べれば、
比べ物にならない体付きだろう。
トーレスはユーロ決勝の試合後にも自然と各選手の子供達に取り囲まれたいた。
優男かどうかは体付きよりも顔立ちの方が重要だろう。
比べ物にならない体付きだろう。
トーレスはユーロ決勝の試合後にも自然と各選手の子供達に取り囲まれたいた。
優男かどうかは体付きよりも顔立ちの方が重要だろう。
容姿だけの評価なら世界的にはアラブと東欧が最高地域のようですね。
ニーナ・ドブレフ最高
優男志向は昔からある西洋にもあった
筋肉むちむちは最近アメリカだけ
アメリカの文化である
筋肉むちむちは最近アメリカだけ
アメリカの文化である
49 :世界@名無史さん:2012/08/25(土) 00:07:11.15 0
ところで、なぜ日本において、高学歴者は容姿端麗に見えるのでしょうか?
育ち方によって容姿は変わってくるものなのでしょうか?
タン・レイ
http://www.ted.com/talks/lang/ja/tan_le_a_headset_that_reads_your_brainwaves.html
育ち方によって容姿は変わってくるものなのでしょうか?
タン・レイ
http://www.ted.com/talks/lang/ja/tan_le_a_headset_that_reads_your_brainwaves.html
>>18
18世紀19世紀の前半の頃に「黄色人種」というとトルコ人やアラビア人やペルシャ人のことだったから・・・・・・
18世紀19世紀の前半の頃に「黄色人種」というとトルコ人やアラビア人やペルシャ人のことだったから・・・・・・
51 :名無し募集中。。。:2012/08/25(土) 10:19:20.43 0
黄色人種ってモンゴロイドの意味だろ
>>48
そうだったのか。
そうだったのか。
とは言え、ヨーロッパの美男子…アランドロンとかジェームス・ボンド役にしても極端にムキムキではないにしても
それなりに筋肉質じゃないかと思うのだが。
それなりに筋肉質じゃないかと思うのだが。
54 :世界@名無史さん:2012/08/26(日) 01:27:34.83 0
日本のアニメキャラクターは白人なのでしょうか?
「日本のアニメキャラクター」としかいいようのない何かだ。
それに金髪とか白人っぽい記号的属性がついてたりするだけ。
それに金髪とか白人っぽい記号的属性がついてたりするだけ。
http://chirashi-archive.com/img/20110415-14a.jpg
http://farm3.static.flickr.com/2591/5843125998_401ddc5286_b.jpg
日本は美しい…日本は素晴らしい……だけど日本人は…という服飾業界の嫌味か?
{kind=link}
http://farm3.static.flickr.com/2591/5843125998_401ddc5286_b.jpg
{kind=link}
日本は美しい…日本は素晴らしい……だけど日本人は…という服飾業界の嫌味か?
ファッション誌を見ても分かるが、
普通の日本人の体型に合わないようなファッションや
パリの最新の流行がとか、
だれが一番の戦犯なんだろw
普通の日本人の体型に合わないようなファッションや
パリの最新の流行がとか、
だれが一番の戦犯なんだろw
世界的に問題となっている人種差別は、歴史的にはある時期から白人が富と力を得てから発生したものです。
ヨーロッパやアメリカで大きな社会問題となっている人種差別の根源は、全て白人問題であると言われています。
ヨーロッパやアメリカで大きな社会問題となっている人種差別の根源は、全て白人問題であると言われています。
61 :名無し募集中。。。:2012/08/26(日) 11:34:02.65 0
>>58
これはw
これはw
62 :名無し募集中。。。:2012/08/26(日) 11:34:41.77 0
エジプトの歴史でも、エジプト南部の黒人が北上して、エジプトを支配した時代があったような…。
当時のエジプトでは、古代エジプトの文化や伝統が失われていて、むしろその黒人支配層が
その文化を保持していたから、逆に文化復興になったんだよな。
その時代だと、アラブ<<黒人なのかなw
当時のエジプトでは、古代エジプトの文化や伝統が失われていて、むしろその黒人支配層が
その文化を保持していたから、逆に文化復興になったんだよな。
その時代だと、アラブ<<黒人なのかなw
エジプトは黒人だろ
65 :名無し募集中。。。:2012/08/26(日) 17:25:18.04 0
サハラ砂漠より北がコーカソイドのアラブ人
南がいわゆる奴隷黒人になる
南がいわゆる奴隷黒人になる
それは新しい時代のことだろ。
イスラム帝国でアラビア人が流入、その後オスマン帝国が東欧から兵員や農耕奴隷をどっさり投入して、
すっかり白くなった。
イベリアを700年間支配したモール人なんて真っ黒けだったし。その前の古代のリビア人やらエジプト人も
肌の黒い人だったし。
イスラム帝国でアラビア人が流入、その後オスマン帝国が東欧から兵員や農耕奴隷をどっさり投入して、
すっかり白くなった。
イベリアを700年間支配したモール人なんて真っ黒けだったし。その前の古代のリビア人やらエジプト人も
肌の黒い人だったし。
人種的にはコーカソイドなのは?ちょい肌の色は黒いのだろうけどね。
ムーア人が黒人だ という観念も、シェークスピアの戯曲『オセロ』によって誤って広められたものだそうで。
ムーア人が黒人だ という観念も、シェークスピアの戯曲『オセロ』によって誤って広められたものだそうで。
エジプトが黒人とすると、世界最初の文字も黒人で、世界最初の文明も黒人ということに…w
(メソポタミアのモノより、500年ほど前の文字が見つかっているんだよな)
今のアルファベットも元を正せば、エジプトのヒエログリフな訳で。
黒人が発明したものをつかっているということに…。
まあ、人種差別満開で考えるのも良いけどさーw
(メソポタミアのモノより、500年ほど前の文字が見つかっているんだよな)
今のアルファベットも元を正せば、エジプトのヒエログリフな訳で。
黒人が発明したものをつかっているということに…。
まあ、人種差別満開で考えるのも良いけどさーw
肌の白い黒いは創造性や知能にあまり関係ないというけれど・・・・・・
70 :名無し募集中。。。:2012/08/26(日) 22:15:19.13 0
71 :名無し募集中。。。:2012/08/26(日) 22:16:01.32 0
シェークスピアってムハンマドとアリーが地獄に落ちてるってやつだろw
72 :名無し募集中。。。:2012/08/26(日) 22:17:02.53 0
>>72
単に産業革命おこして、地球を制覇しちゃっただけだろ。
単に産業革命おこして、地球を制覇しちゃっただけだろ。
ま知能は遺伝らしいけどな
おまえらあきらめろ
おまえらあきらめろ
IQ検査したら、 東洋人>西洋人 だったじゃないか。
完全に遺伝子が一致する双子の研究では、運動系や美術系の成績は一番遺伝子の影響があるそうな。
でも、数学は一番遺伝子の影響が少ないとか。
完全に遺伝子が一致する双子の研究では、運動系や美術系の成績は一番遺伝子の影響があるそうな。
でも、数学は一番遺伝子の影響が少ないとか。
>>66
古代エジプト人はイスラエル人やフェニキア人と同じセム系では?
古代エジプト人はイスラエル人やフェニキア人と同じセム系では?
>>76
15王朝期の支配層はセム系だといわれているね
15王朝期の支配層はセム系だといわれているね
http://jisaku.155cm.com/src/1341284504_51bf0a84822b2d01eacb4ab285a56c6eaac94de4.jpg
http://jisaku.155cm.com/src/1341284504_d2257e9515e28bf4c86b887d8de1f697a107244e.jpg
http://jisaku.155cm.com/src/1341284504_86dbbf87227ef75ba3d9ccd90c92f41eb8136a96.jpg
http://jisaku.155cm.com/src/1341284504_7afd14343e3f7c722811a6242584c6c6523843f7.jpg
http://jisaku.155cm.com/src/1341284504_dd480694f47972e2b22aa13c62a78fa5b323cfef.jpg
http://jisaku.155cm.com/src/1341284504_6d6c82adaf7e644249863d0fb112980d2c303d03.jpg
http://jisaku.155cm.com/src/1341284504_dd408a584cc44c0e4f1aee99db3af586d0caf280.jpg
http://jisaku.155cm.com/src/1341284504_95bccb69b179ad2a78f0765ced3f49ce806db5eb.jpg
http://jisaku.155cm.com/src/1341284504_9f378276beb83090637765f002506a13de3b88ae.jpg
{kind=link}
http://jisaku.155cm.com/src/1341284504_d2257e9515e28bf4c86b887d8de1f697a107244e.jpg
{kind=link}
http://jisaku.155cm.com/src/1341284504_86dbbf87227ef75ba3d9ccd90c92f41eb8136a96.jpg
{kind=link}
http://jisaku.155cm.com/src/1341284504_7afd14343e3f7c722811a6242584c6c6523843f7.jpg
{kind=link}
http://jisaku.155cm.com/src/1341284504_dd480694f47972e2b22aa13c62a78fa5b323cfef.jpg
{kind=link}
http://jisaku.155cm.com/src/1341284504_6d6c82adaf7e644249863d0fb112980d2c303d03.jpg
{kind=link}
http://jisaku.155cm.com/src/1341284504_dd408a584cc44c0e4f1aee99db3af586d0caf280.jpg
{kind=link}
http://jisaku.155cm.com/src/1341284504_95bccb69b179ad2a78f0765ced3f49ce806db5eb.jpg
{kind=link}
http://jisaku.155cm.com/src/1341284504_9f378276beb83090637765f002506a13de3b88ae.jpg
{kind=link}
>>77
それ以前のも、セム系なんじゃないの?
それ以前のも、セム系なんじゃないの?
ていうか>>66の「エジプトにオスマン帝国が東欧から兵員や農耕奴隷をどっさり投入」って
出典は? 思いつきで書いてない?
オスマン帝国がマムルーク朝を征服した後、総督なんかを置いた後
従来のマムルーク体制をほとんどそのまま残して税だけ取り立ててたはずだけど
出典は? 思いつきで書いてない?
オスマン帝国がマムルーク朝を征服した後、総督なんかを置いた後
従来のマムルーク体制をほとんどそのまま残して税だけ取り立ててたはずだけど
81 :名無し募集中。。。:2012/08/28(火) 06:48:06.07 0
82 :名無し募集中。。。:2012/08/28(火) 06:48:43.55 0
15世紀にキリスト教徒の子弟から優秀な青少年を徴集し、
イスラム教に改宗させてイェニチェリなどに採用する
デヴシルメ制度が考案され、定期的な人材供給が行われるようになる。
イスラム教に改宗させてイェニチェリなどに採用する
デヴシルメ制度が考案され、定期的な人材供給が行われるようになる。
>>81-82
デヴシルメが行われていたのはルーメリアとアナトリア。
エジプトではマムルーク軍人の勢力が18世紀にムハンマド・アリーに虐殺されるまで温存された。
だいたいイェニチェリを農民として植民させてどうするよ。しかも中東では元々人口の多いエジプトにw
デヴシルメが行われていたのはルーメリアとアナトリア。
エジプトではマムルーク軍人の勢力が18世紀にムハンマド・アリーに虐殺されるまで温存された。
だいたいイェニチェリを農民として植民させてどうするよ。しかも中東では元々人口の多いエジプトにw
84 :世界@名無史さん:2012/08/28(火) 16:32:29.96 0
「ポーンズ・イン・ザ・ゲーム」ウィリアム・ギー・カー著
・アルコール飲料、麻薬、道徳的退廃、その他全ての悪を大衆の中に注入せよ
・自由、平等、博愛のスローガンで大衆を欺瞞せよ
・プロパガンダを行え
・青年、若者を虜にする事が重要
(「豚」にはサッカー野球パチンコ等の「玉転がし」を与えよ)
85 :世界@名無史さん:2012/08/28(火) 22:51:52.43 0
玄葉「韓国は都合が悪いことあると電話に出ない!居留守使うんです」
ttp://www.youtube.com/watch?v=xInlLlGMIO8&feature=g-logo-xre
ttp://www.youtube.com/watch?v=xInlLlGMIO8&feature=g-logo-xre
NHKがきたら、居留守する国があるんだけど
87 :名無し募集中。。。:2012/08/29(水) 03:26:08.75 0
>>83
そもそもマムルークそのものが白人系軍事奴隷だからね
ロシア南部あたりの遊牧民を奴隷として買ってきたんだから
昔からイスラム世界には白人系軍事奴隷がたくさんいたと思う
バイバルスだってスラブ系でしょ
そもそもマムルークそのものが白人系軍事奴隷だからね
ロシア南部あたりの遊牧民を奴隷として買ってきたんだから
昔からイスラム世界には白人系軍事奴隷がたくさんいたと思う
バイバルスだってスラブ系でしょ
>>22
金髪碧眼、色白、高身長で手足が長くマネキンの様な風貌、スタイルはゲルマン系
北欧が一番綺麗だろあとスラブ系 地中海人種は元々はチュニジア(カルタゴ)人
と同じ人種 欧州はゲルマン民族の大移動でゲルマンの血が欧州全体にばら撒かれた
イベリア半島(西ゴート族)フランス(フランク族)イングランド(アングロサクソン族)
欧州中央(ブルグント族)北イタリア(ロンバルト族)(東ゴート族)など ブロンドで青い目がゲルマンの血
金髪碧眼、色白、高身長で手足が長くマネキンの様な風貌、スタイルはゲルマン系
北欧が一番綺麗だろあとスラブ系 地中海人種は元々はチュニジア(カルタゴ)人
と同じ人種 欧州はゲルマン民族の大移動でゲルマンの血が欧州全体にばら撒かれた
イベリア半島(西ゴート族)フランス(フランク族)イングランド(アングロサクソン族)
欧州中央(ブルグント族)北イタリア(ロンバルト族)(東ゴート族)など ブロンドで青い目がゲルマンの血
>>87
中世イスラム圏で白人奴隷というのは、黒人やドラヴィダ系インド人(ハムの子孫と総称される)
ではないそれ以外の連中という意味で、アジア系遊牧民等も含まれる。
バイバルスの頃のマムルークはスラヴではなくテュルク系のキプチャク族。
中世イスラム圏で白人奴隷というのは、黒人やドラヴィダ系インド人(ハムの子孫と総称される)
ではないそれ以外の連中という意味で、アジア系遊牧民等も含まれる。
バイバルスの頃のマムルークはスラヴではなくテュルク系のキプチャク族。
91 :世界@名無史さん:2012/08/29(水) 13:31:32.97 0
>>75
容姿端麗か否かは客観的で科学的な数値にして図れない。
しかし、頭の良し悪しは数値にして図れる(IQ検査など)。
知的障害者を診断する際の公的な基準になっているので、客観的で科学的な数値である。
科学の発展によって、今では発達障害者という新たな枠組みも公のものになっている。
大東亜共栄圏を再び行うならば、東亜人のIQをことさら取り上げてやるとよいだろう。
容姿端麗か否かは客観的で科学的な数値にして図れない。
しかし、頭の良し悪しは数値にして図れる(IQ検査など)。
知的障害者を診断する際の公的な基準になっているので、客観的で科学的な数値である。
科学の発展によって、今では発達障害者という新たな枠組みも公のものになっている。
大東亜共栄圏を再び行うならば、東亜人のIQをことさら取り上げてやるとよいだろう。
つーかさ
新モンゴロイドも北欧系コーカソイドも同じ寒冷地への適応のはずなのに真逆の変化
人類学っていい加減だよな
新モンゴロイドも北欧系コーカソイドも同じ寒冷地への適応のはずなのに真逆の変化
人類学っていい加減だよな
サルが進化したら、必ず人間になるとは限らないんだよ。たとえ同じ刺激であってもね。
95 :名無し募集中。。。:2012/08/30(木) 09:22:08.32 0
>>95
ロシア人に征服されて混血しまくってる今のタタール人と13世紀のキプチャク族が見た目一緒だとする根拠は?
ロシアの支配下に入るのが遅かったカザフスタンあたりだと日本で歩いてても違和感ないのがいっぱいいるw
ttp://www.youtube.com/watch?v=Z-iY7JUUWC4
ロシア人に征服されて混血しまくってる今のタタール人と13世紀のキプチャク族が見た目一緒だとする根拠は?
ロシアの支配下に入るのが遅かったカザフスタンあたりだと日本で歩いてても違和感ないのがいっぱいいるw
ttp://www.youtube.com/watch?v=Z-iY7JUUWC4
テュルク系の人間っていつからコーカソイド顔になったん
98 :名無し募集中。。。:2012/08/30(木) 20:11:18.10 0
99 :世界@名無史さん:2012/08/30(木) 21:27:20.49 0
>>96
ウィグル始めチュルク系はアーリア人のR1a遺伝子が多いからコーカソイド系の人間がチュルク語化したと思う。
ウィグル始めチュルク系はアーリア人のR1a遺伝子が多いからコーカソイド系の人間がチュルク語化したと思う。
Q.トルコ人は何故あれ程広大な大帝国を建設したのか?
A.トルコ人の女性はあまりにも醜く、トルコの男が多民族の女性を渇望していたからである
by レオポルト・フォン・ランケ(ドイツ人・「近代歴史学の父」)
A.トルコ人の女性はあまりにも醜く、トルコの男が多民族の女性を渇望していたからである
by レオポルト・フォン・ランケ(ドイツ人・「近代歴史学の父」)
101 :名無し募集中。。。:2012/08/30(木) 21:51:52.10 0
ギリシア独立運動
オスマン帝国
アーリア人至上主義
ギリシア彫刻
オスマン帝国
アーリア人至上主義
ギリシア彫刻
102 :名無し募集中。。。:2012/08/30(木) 21:53:53.68 0
>>100
>レオポルト・フォン・ランケ
オスマン帝国末期にギリシアが独立したわけで
そういう歴史をふまえて当時の人間を理解しないといけないのよ
ヨーロッパの起源をギリシア・ローマだと考えていたヨーロッパ人が
ギリシアを支配していたトルコ人を悪くいうのは当たり前
>レオポルト・フォン・ランケ
オスマン帝国末期にギリシアが独立したわけで
そういう歴史をふまえて当時の人間を理解しないといけないのよ
ヨーロッパの起源をギリシア・ローマだと考えていたヨーロッパ人が
ギリシアを支配していたトルコ人を悪くいうのは当たり前
103 :名無し募集中。。。:2012/08/30(木) 21:54:53.14 0
104 :世界@名無史さん:2012/08/31(金) 02:45:21.44 0
ヨーロッパの起源って何処さ?
105 :名無し募集中。。。:2012/08/31(金) 07:03:21.94 0
ヨーロッパの起源は古代ローマ帝国でしょ
107 :世界@名無史さん:2012/08/31(金) 13:43:46.92 0
>>92
世界一のイケメンはLed zeppelinのロバート・プラントじゃね?
世界一のイケメンはLed zeppelinのロバート・プラントじゃね?
108 :名無し募集中。。。:2012/08/31(金) 13:54:00.09 0
女優はMélanie Laurentが美人だと思う
背は高くないけど
背は高くないけど
>>104
元々のオリエント(Orient)の語源とはラテン語の「太陽の昇るところ」オリエンス(Oriens)に由来し、
古代のイタリア半島から見て「東方」はギリシアを指していた。それが後、極東を含めたアジア全域を指すようになった。
ヨーロッパ(Europe)という呼称は、ギリシア語のエレボス(erebus)闇という語源。そんなギリシア人から見た
昼でも暗く深い森林に覆われた黒い森の住人のイメージから来ている。
元々のオリエント(Orient)の語源とはラテン語の「太陽の昇るところ」オリエンス(Oriens)に由来し、
古代のイタリア半島から見て「東方」はギリシアを指していた。それが後、極東を含めたアジア全域を指すようになった。
ヨーロッパ(Europe)という呼称は、ギリシア語のエレボス(erebus)闇という語源。そんなギリシア人から見た
昼でも暗く深い森林に覆われた黒い森の住人のイメージから来ている。
>1
の例から見ても分かるように美的感覚は時代における権力の推移に付随しているということが観察される。
の例から見ても分かるように美的感覚は時代における権力の推移に付随しているということが観察される。
111 :世界@名無史さん:2012/09/02(日) 12:45:25.49 0
パルテノン神殿(イメージ) イメージ!イメージ!イメージダウン!
近代欧州は古代ギリシャへの憧れが強く、20世紀初頭の英国では本物のギリシャ彫刻が彩色されていても
ギリシャ彫刻は白いというイメージがあり、イメージ通りにする為にあえて脱色するような事までやってしまったらしい。
近代欧州は古代ギリシャへの憧れが強く、20世紀初頭の英国では本物のギリシャ彫刻が彩色されていても
ギリシャ彫刻は白いというイメージがあり、イメージ通りにする為にあえて脱色するような事までやってしまったらしい。
兵馬俑もそうだな。本当はカラフルに彩色されてたのに
遺跡を開けた途端新鮮な空気に触れて脱色してしまったらしい。
遺跡を開けた途端新鮮な空気に触れて脱色してしまったらしい。
日本の寺院仏閣も建築当初はカラフルな原色ギラギラだったと言うしな
奈良の大仏の建造当初の復元図がまんま大阪のオバちゃんカラーだったのには驚いた
奈良の大仏の建造当初の復元図がまんま大阪のオバちゃんカラーだったのには驚いた
ドイツの映画みてるとほんとに美人が出てこないと思う
でも、まれに金髪で青い大きな目の華奢な美少女が出てくる時があって
最後の字幕で名前チェックしたら、ヴェラとかナジェジダとかリュドミラとか
なんとかスカヤみたいな名前だったりする
でも、まれに金髪で青い大きな目の華奢な美少女が出てくる時があって
最後の字幕で名前チェックしたら、ヴェラとかナジェジダとかリュドミラとか
なんとかスカヤみたいな名前だったりする
それ面白いなw ナジェージダは間違いなくスラヴ系の名前。
116 :世界@名無史さん:2012/09/14(金) 20:46:43.88 0
ヒトラー最後の12日間で秘書ユンゲの役やってた女優もルーマニア人でした
おかしいと思ったんだよ、ドイツ人でこんな綺麗どころとか
おかしいと思ったんだよ、ドイツ人でこんな綺麗どころとか
117 :世界@名無史さん:2012/09/14(金) 22:31:15.14 0
↑
少女兵もロシア人だった。
ヒトラーはいわゆるテーブル・トークで「なぜウクライナの少年少女はあれほど美しいのに、大人になると醜くなってしまうのだろうか?」というような事を言って、嘆いていた。
ナチスって東欧から子供を拉致してドイツ人として育てることをしていなかったか?
もしかすると、過去から現在に至るまで、形を変えつつドイツ人の美的センスは受け継がれているのかもしれない。
話は変わって、フィリピン人の英会話講師と話をしていたが、
フィリピンのCMの役者は殆どが欧米人だとのこと。
これは、フィリピンが米文化の影響を強く受けているからか、それとも欧米人の顔がより「普遍的」で「世界的」だからか。
台湾では日本がステータスで日本語や日本のキャラクターが受けているが、日本人の顔が受けているという話は聞いたことがない。
個人的にはアメリカと軍事的に、あるいは経済的にむすびつきが強くなっているベトナムの広告がどのように変わっていくかに興味がある。
少女兵もロシア人だった。
ヒトラーはいわゆるテーブル・トークで「なぜウクライナの少年少女はあれほど美しいのに、大人になると醜くなってしまうのだろうか?」というような事を言って、嘆いていた。
ナチスって東欧から子供を拉致してドイツ人として育てることをしていなかったか?
もしかすると、過去から現在に至るまで、形を変えつつドイツ人の美的センスは受け継がれているのかもしれない。
話は変わって、フィリピン人の英会話講師と話をしていたが、
フィリピンのCMの役者は殆どが欧米人だとのこと。
これは、フィリピンが米文化の影響を強く受けているからか、それとも欧米人の顔がより「普遍的」で「世界的」だからか。
台湾では日本がステータスで日本語や日本のキャラクターが受けているが、日本人の顔が受けているという話は聞いたことがない。
個人的にはアメリカと軍事的に、あるいは経済的にむすびつきが強くなっているベトナムの広告がどのように変わっていくかに興味がある。
118 :世界@名無史さん:2012/09/14(金) 23:19:01.00 0
ベトナムはフランスとの結びつきも忘れないでほしいなあ
ドヌーヴ主演の『インドシナ』を観てからその思いを強くした
ドヌーヴ主演の『インドシナ』を観てからその思いを強くした
マレーネ・ディートリヒは例外。あれをドイツ女性一般と考えてはいけない。
スケートや体操や新体操見ると、美人や美少女が多いよね
ベレズナヤ ナフカ様 ドミトリエワ カナエワ様 サフチェンコ リアシェンコ
デンコワ コモワ トゥクタミシェワ コロベイニコワ シェレペン リヤザノワ
ブットゥイルスカヤ姐 マリニナ ホフロワ ベッソノワ ゲルボルト バザロワ
ドイツ、オランダ、北欧あたりはその楽しみが少ない
派手な金髪と青い小さな眼とピンクの肌 ごつい体格 地味な雰囲気
金髪とブルーアイズが似合わない人が多いような
個人的な好みだけどね
>>119
ナスターシャ・キンスキーがいるよ
でもあの人は…
ベレズナヤ ナフカ様 ドミトリエワ カナエワ様 サフチェンコ リアシェンコ
デンコワ コモワ トゥクタミシェワ コロベイニコワ シェレペン リヤザノワ
ブットゥイルスカヤ姐 マリニナ ホフロワ ベッソノワ ゲルボルト バザロワ
ドイツ、オランダ、北欧あたりはその楽しみが少ない
派手な金髪と青い小さな眼とピンクの肌 ごつい体格 地味な雰囲気
金髪とブルーアイズが似合わない人が多いような
個人的な好みだけどね
>>119
ナスターシャ・キンスキーがいるよ
でもあの人は…
{kind=link}
ごっついけど綺麗なブリジット・ニールセン(20年前)萌えな漏れ
124 :世界@名無史さん:2012/09/16(日) 20:58:45.00 0
当然だが、ドイツ人は純粋な人種ではない。
スラブ系の血が入っている。
ヒトラーもチェコ人の末裔である。
逆に、他人種の血が入って不細工になった民族っているの?
スラブ系の血が入っている。
ヒトラーもチェコ人の末裔である。
逆に、他人種の血が入って不細工になった民族っているの?
チェコ人と北欧人ならチェコ人のが美しいと思う。
モンゴル人と混血した民族
整形すれば無問題
ソフィア・ローレンの個性的顔は地中海人種の典型
ギリシア彫刻のモデルは古代ゲルマン民族
あの辺なら東ゴート人 ギリシア彫刻は今は白だが、彩色されていた。
ブロンドのヘアーに青い目という金髪碧眼 白い肌のあまりの美しさに
当時のギリシア人が形に残そうとして・・・ミロのビーナスも金髪碧眼で
均整のとれた眩しいばかりの白い肌だった。当時のギリシア人にない高身長で
均整のとれた美しい裸体は当時のギリシア人にとって憧れだった。
あの辺なら東ゴート人 ギリシア彫刻は今は白だが、彩色されていた。
ブロンドのヘアーに青い目という金髪碧眼 白い肌のあまりの美しさに
当時のギリシア人が形に残そうとして・・・ミロのビーナスも金髪碧眼で
均整のとれた眩しいばかりの白い肌だった。当時のギリシア人にない高身長で
均整のとれた美しい裸体は当時のギリシア人にとって憧れだった。
エレボスの闇から突然現れた眩しいばかりの美しさと讃えている
エレボスの闇とは当時のヨーロッパの深く黒い森の例え
エレボスの闇とは当時のヨーロッパの深く黒い森の例え
131 :世界@名無史さん:2012/09/18(火) 14:46:12.07 0
アングロサクソンの容姿が端麗ってことになったのは
アングロサクソンが世界の軍事・経済・情報の覇権をとったからだろ
アングロサクソンが世界の軍事・経済・情報の覇権をとったからだろ
いや
イギリスが改宗する伝説にアングロサクソンの容姿をローマ法王が褒める逸話があったな
イギリスが改宗する伝説にアングロサクソンの容姿をローマ法王が褒める逸話があったな
133 :世界@名無史さん:2012/09/18(火) 17:48:51.63 0
クレゴリウス1世大教皇がローマ市街で金髪の美少年奴隷を褒めたやつね
アングル族の奴隷と聞いて「なんと!北にはエンジェル族がいるのか?」って聞き間違えた・・・
とかいう尾ヒレが付いていた
アングル族の奴隷と聞いて「なんと!北にはエンジェル族がいるのか?」って聞き間違えた・・・
とかいう尾ヒレが付いていた
>>132-133
検索してもこの板しかヒットしないんだけどw
検索してもこの板しかヒットしないんだけどw
大体、モンゴル人に支配されていたロシアではモンゴル人的要素を持った人たちを美しいとしたしな。
現在でも…このスレの過去ログでも、ロシア人は東洋、西洋にあまりこだわらないという報告あるよね。
現在でも…このスレの過去ログでも、ロシア人は東洋、西洋にあまりこだわらないという報告あるよね。
136 :世界@名無史さん:2012/09/18(火) 19:14:02.95 0
>>136
そうなのか。まあ、確かにエンジェルの西洋画ってイメージとして、金髪で巻髪の幼児の容貌だしなあ。
そうなのか。まあ、確かにエンジェルの西洋画ってイメージとして、金髪で巻髪の幼児の容貌だしなあ。
138 :世界@名無史さん:2012/09/18(火) 21:09:28.47 0
>>135
ロシアやウクライナはアジア人っぽい顔が平べったい白人も多いけど、
トップは北欧人などノルマン系を遥かに凌駕する凄まじい美形だよな。
この人とかすげー好みなんだけど↓
http://youtu.be/sWIxuLz5SGs?t=20s
北欧人はなんか表情が冷たすぎるし金髪に碧眼だと人間じゃないみたいなんだよな。本当映画や漫画に出てくるエルフのような別の種族に見えてくる。
ブルネットに碧眼の白人が一番いいよ
ロシアやウクライナはアジア人っぽい顔が平べったい白人も多いけど、
トップは北欧人などノルマン系を遥かに凌駕する凄まじい美形だよな。
この人とかすげー好みなんだけど↓
http://youtu.be/sWIxuLz5SGs?t=20s
北欧人はなんか表情が冷たすぎるし金髪に碧眼だと人間じゃないみたいなんだよな。本当映画や漫画に出てくるエルフのような別の種族に見えてくる。
ブルネットに碧眼の白人が一番いいよ
>>138
んー。やはり個人の好みの差が…ww
んー。やはり個人の好みの差が…ww
>>132 >>133
その手の話はプロパガンダ的な意味あいで
アングロ・サクソン側が作って広めた可能性がある
原典にあたらないと何とも判断できない
江戸末期開国したときに西洋人をはじめて見た日本人が
「なんて醜い人々だろう」と同情したって逸話を見たんだが
先入観さえなければ異民族の容姿なんてそんなもんじゃないかね
>>138 ストックホルムに3か月ぐらいいたけど
別の種族に見えるようなイケメンや美女なんていなかったぜ・・・
その手の話はプロパガンダ的な意味あいで
アングロ・サクソン側が作って広めた可能性がある
原典にあたらないと何とも判断できない
江戸末期開国したときに西洋人をはじめて見た日本人が
「なんて醜い人々だろう」と同情したって逸話を見たんだが
先入観さえなければ異民族の容姿なんてそんなもんじゃないかね
>>138 ストックホルムに3か月ぐらいいたけど
別の種族に見えるようなイケメンや美女なんていなかったぜ・・・
141 :世界@名無史さん:2012/09/19(水) 19:09:37.57 0
きれいだし美人だとは思うけど
エルフのように美しいとか別の生き物のように美しいかといえばなんか違う
留学先や仕事先の海外で出会った美人のお姉さんって印象
好みでいえばこれくらいなら日本のそのへんの美人のほうがいい
それに異民族ならラテンが一番美人だと思ってるし
エルフのように美しいとか別の生き物のように美しいかといえばなんか違う
留学先や仕事先の海外で出会った美人のお姉さんって印象
好みでいえばこれくらいなら日本のそのへんの美人のほうがいい
それに異民族ならラテンが一番美人だと思ってるし
>>140
初出はベーダの教会史のようだね
初出はベーダの教会史のようだね
しかしアニメで育った日本人の感覚からすると鼻のでかさがアウト
145 :世界@名無史さん:2012/09/23(日) 20:14:53.88 0
ただでさえアジアに比べて人口が少ないのに、ペストで人口の3分2が死んで、さらに領主の初夜権やら地域で凝り固まって血が濃い筈なのになんで白人は美人が多いんだろ?
血が濃くなると絶対ブサになる筈なのにな(例 半島や島国の日本)
アジアに比べて不利な条件だらけなのになー
血が濃くなると絶対ブサになる筈なのにな(例 半島や島国の日本)
アジアに比べて不利な条件だらけなのになー
*賞味期限は20歳までです
147 :世界@名無史さん:2012/09/24(月) 01:05:16.04 0
そうかな。白人のおばさんって結構綺麗な人いるよ。
148 :世界@名無史さん:2012/09/24(月) 09:22:48.54 0
イギリスの征服王の顔は「日本人」
http://ichiba.geocities.jp/gbsg0309/0204/49/316.html
関ヶ原の戦いは、この征服王を記念して行われたことは、間違いない。
149 :名無し募集中。。。:2012/09/24(月) 14:32:57.22 0
メラニーロランが一番美人だと思う
150 :世界@名無史さん:2012/09/24(月) 15:34:08.28 0
>>149
その人アシュケナジーユダヤ人だよ。
その人アシュケナジーユダヤ人だよ。
152 :世界@名無史さん:2012/09/25(火) 19:26:38.22 0
そこらかしこでセックスしまくってたから血が混ざり合って美形になったんじゃないのか
アジア人って現代で見ても同じ民族同士で凝り固まる習性があるというか、行動的じゃないよね。白人と比べて
アジア人って現代で見ても同じ民族同士で凝り固まる習性があるというか、行動的じゃないよね。白人と比べて
KKKの悪口はそこまでだ
品種が違うからだよ
容姿端麗
http://upload.wikimedia.org/wikipedia/commons/thumb/4/4f/Berger_allemand_en_montagne.jpg/800px-Berger_allemand_en_montagne.jpg
淘汰予備軍
http://shinjyuku-seitai.jp/wp-content/themes/keniwp_brown03_g12r/images/twiphoto/600-450-9-25-08.jpg
容姿端麗
http://upload.wikimedia.org/wikipedia/commons/thumb/4/4f/Berger_allemand_en_montagne.jpg/800px-Berger_allemand_en_montagne.jpg
{kind=link}
淘汰予備軍
http://shinjyuku-seitai.jp/wp-content/themes/keniwp_brown03_g12r/images/twiphoto/600-450-9-25-08.jpg
{kind=link}
155 :世界@名無史さん:2012/09/25(火) 22:28:27.14 0
中世ヨーロッパの貴族の間では、小さい足はステータスだったらしいけど(デカい足は労働者階級として忌み嫌われた?)
今でもそういう傾向あるのかな?背は高くても足は小さいほうがいいとか?
今でもそういう傾向あるのかな?背は高くても足は小さいほうがいいとか?
整備されたトラックじゃなく、ジャングルや、藪の森を踏破するような競技が一般的なら、
身長が低い方が有利でモテたろうなw
身長が低い方が有利でモテたろうなw
>>152
スイスを例にあげると
村と村で文化も行政も違うし今でも固まってるよ
地形的に移動が難しいとアジアとかヨーロッパとか関係なくそうなる
逆にハンガリーとかあのへんは地形的に移動がしやすいので
侵入侵略されやすくて対オスマントルコの前線になった
だいたいアジアっつったって日本と大陸、大陸だって中央アジアと沿岸部で
民族も人種も違うのにアジア人でひとくくりにするのはおかしい
スイスを例にあげると
村と村で文化も行政も違うし今でも固まってるよ
地形的に移動が難しいとアジアとかヨーロッパとか関係なくそうなる
逆にハンガリーとかあのへんは地形的に移動がしやすいので
侵入侵略されやすくて対オスマントルコの前線になった
だいたいアジアっつったって日本と大陸、大陸だって中央アジアと沿岸部で
民族も人種も違うのにアジア人でひとくくりにするのはおかしい
ハンガリーはカルパチア山脈の向こう側のバルカン半島に比べればかなり平和だぞw
トルコとハプスブルクの前線になったのはスレイマン大帝がヤゲロー朝を滅ぼして
ハンガリーを分割してから。
トルコとハプスブルクの前線になったのはスレイマン大帝がヤゲロー朝を滅ぼして
ハンガリーを分割してから。
ハンガリーって平坦でだだっ広いんだぞ
あのへんになったら急に視界が開けた
そりゃこんな平坦だったら往来もしやすいわと思った
あのへんになったら急に視界が開けた
そりゃこんな平坦だったら往来もしやすいわと思った
160 :世界@名無史さん:2012/09/27(木) 19:11:57.68 0
ヨーロッパでもハンガリーは美人多いよな。チェコスロバキアハンガリーロシアこの3強がヤバイ。
女子柔道のヘドヴィグ・カラカス美人
162 :世界@名無史さん:2012/10/01(月) 23:05:46.56 0
ヨーロッパの絵画とか見るとブスなんだよな不思議
だからそれが美的感覚の違い。
164 :世界@名無史さん:2012/10/06(土) 18:51:33.18 0
ヨーロッパ人に美人の多い理由
古代からラテン系、ケルト系、ゲルマン系が入り乱れ混血しまくったから
これらをベースに加えてノルマン系、フン族、マジャール、スラブ系がブレンドされたのがヨーロッパ人
アジアで例えるなら縄文人、漢民族を初めとする弥生系(日中韓モンゴル系)、東南アジアン、アイヌがブレンドされまくったようなもん。
あの広い中国大陸でさえヨーロッパに比べて全然混血してないから不細工が多い
古代からラテン系、ケルト系、ゲルマン系が入り乱れ混血しまくったから
これらをベースに加えてノルマン系、フン族、マジャール、スラブ系がブレンドされたのがヨーロッパ人
アジアで例えるなら縄文人、漢民族を初めとする弥生系(日中韓モンゴル系)、東南アジアン、アイヌがブレンドされまくったようなもん。
あの広い中国大陸でさえヨーロッパに比べて全然混血してないから不細工が多い
混血したら美形になると理由ならば、
世界各地の動物は内陸ほど美形になるということだが、
実際はホッキョクグマとヒグマの容姿に大差ない。
白人は眼が大きいから美形なだけだと思う。
美形に意味は全然無いと思う。
世界各地の動物は内陸ほど美形になるということだが、
実際はホッキョクグマとヒグマの容姿に大差ない。
白人は眼が大きいから美形なだけだと思う。
美形に意味は全然無いと思う。
166 :世界@名無史さん:2012/10/06(土) 19:05:49.07 0
>>165
そんなのおかしい。
白人はもう、輪郭からきれいなんだよ。
東洋系は余白が多すぎる。
輪郭もいびつ。
白人は、輪郭も美しく、
さらに目鼻立ちが整い、無駄がない。
顔に無駄がないんだよ。
http://www.allposters.co.jp/-sp/%E3%83%AA%E3%83%B4%E3%83%BB%E3%82%BF%E3%82%A4%E3%83%A9%E3%83%BC-Posters_i991243_.htm
見てみろこれ!!!
一切無駄のない白人特有の顔。
アジア系は丸顔だらけで余白が多く、ろくなもんじゃないw
そんなのおかしい。
白人はもう、輪郭からきれいなんだよ。
東洋系は余白が多すぎる。
輪郭もいびつ。
白人は、輪郭も美しく、
さらに目鼻立ちが整い、無駄がない。
顔に無駄がないんだよ。
http://www.allposters.co.jp/-sp/%E3%83%AA%E3%83%B4%E3%83%BB%E3%82%BF%E3%82%A4%E3%83%A9%E3%83%BC-Posters_i991243_.htm
見てみろこれ!!!
一切無駄のない白人特有の顔。
アジア系は丸顔だらけで余白が多く、ろくなもんじゃないw
芸能人は差し歯とかいろいろやってっからかけらほども基準にならない。
文化的優位の美的感覚への影響を主張する人は、
それ以前にまずこういった外見職業人の容姿を目にする機会が
先進国人の方が圧倒的に多いということを考えるべきだと思う。
後進国人モデルを見れるメディアは無い。
ニュース映像は紛争などの悲惨な民間人が映るのみ。外見が悪いのは当然。
文化的優位の美的感覚への影響を主張する人は、
それ以前にまずこういった外見職業人の容姿を目にする機会が
先進国人の方が圧倒的に多いということを考えるべきだと思う。
後進国人モデルを見れるメディアは無い。
ニュース映像は紛争などの悲惨な民間人が映るのみ。外見が悪いのは当然。
168 :世界@名無史さん:2012/10/06(土) 19:20:15.32 0
>>167
そんなことないだろ。
世界の共通したメディアなんてアメリカくらいだろ。
そのアメリカ文化の象徴たる映画産業。
その中であらゆるコーカソイドとネグロイドは移ってるから、
日本人だってほとんど白人なんて住んでないが、みんな黒人も白人もわかってる。
しかし、あの巨大大国アメリカの映画にすら東洋人の顔を主人公にしたものなどない。
カンフー、忍者、サムライ、など特定のもの意外は絶対、東洋人は移らせてもらえない。
脇役ですらほとんどないのが現実。
世界の共通認識として、東洋人の容姿は論外とされてる。
アメリカのポップミュージックシーンで、白人黒人だらけだが、東洋人アイドルなんて一人も居ないだろ。
アメリカはアジア系は黒人より多くすんでるんだぞ?
そんなことないだろ。
世界の共通したメディアなんてアメリカくらいだろ。
そのアメリカ文化の象徴たる映画産業。
その中であらゆるコーカソイドとネグロイドは移ってるから、
日本人だってほとんど白人なんて住んでないが、みんな黒人も白人もわかってる。
しかし、あの巨大大国アメリカの映画にすら東洋人の顔を主人公にしたものなどない。
カンフー、忍者、サムライ、など特定のもの意外は絶対、東洋人は移らせてもらえない。
脇役ですらほとんどないのが現実。
世界の共通認識として、東洋人の容姿は論外とされてる。
アメリカのポップミュージックシーンで、白人黒人だらけだが、東洋人アイドルなんて一人も居ないだろ。
アメリカはアジア系は黒人より多くすんでるんだぞ?
170 :世界@名無史さん:2012/10/06(土) 21:23:10.76 0
>>168
ナイジェリア人もネグロイドでしょ。
いくらでも類似は映画に出てるでしょ?
でもあれだけアメリカにアジア系住んでても、
東洋系のスターなんて、ジャッキーと忍者だけでしょwって言う意味。
要は需要がないということ。それくらい他人種は東洋系に興味を持たない。といういみ。
ナイジェリア人もネグロイドでしょ。
いくらでも類似は映画に出てるでしょ?
でもあれだけアメリカにアジア系住んでても、
東洋系のスターなんて、ジャッキーと忍者だけでしょwって言う意味。
要は需要がないということ。それくらい他人種は東洋系に興味を持たない。といういみ。
ジュネーヴ協定違反といって良いほどの体臭を放つドラム缶連中
172 :世界@名無史さん:2012/10/06(土) 22:48:52.83 0
173 :世界@名無史さん:2012/10/06(土) 22:52:44.08 0
白人の美形に意味は無い。
猫でもなんでも目玉が適度にでかいと美的感覚に刺激がある。
ただの動物本能。
ではそのことに何の意味もない。
猫は美形でも高等生物になり得ない。
カメラ写りがいいだけ。
映像・画像といった文明の利器がなければ何ももたらさない。無価値。
猫でもなんでも目玉が適度にでかいと美的感覚に刺激がある。
ただの動物本能。
ではそのことに何の意味もない。
猫は美形でも高等生物になり得ない。
カメラ写りがいいだけ。
映像・画像といった文明の利器がなければ何ももたらさない。無価値。
174 :世界@名無史さん:2012/10/06(土) 23:02:45.33 0
>>173
ドイツの歴史見れば、人類最強じゃねいか!!
歴史もすごくて、文明もすごくて、頭もよくて、技術力も凄まじくて、その上美形で......
日本てどんだけドイツ文化ぱくってるかしってるの?
医学もドイツ語 憲法もドイツ 医療保険もドイツ 車はベンツにBM
世界史といえばドイツゲルマン アインシュタインもドイツ生まれ。
言えばきりないほど白人優れてる上での美景ですが何か?
ドイツの歴史見れば、人類最強じゃねいか!!
歴史もすごくて、文明もすごくて、頭もよくて、技術力も凄まじくて、その上美形で......
日本てどんだけドイツ文化ぱくってるかしってるの?
医学もドイツ語 憲法もドイツ 医療保険もドイツ 車はベンツにBM
世界史といえばドイツゲルマン アインシュタインもドイツ生まれ。
言えばきりないほど白人優れてる上での美景ですが何か?
ゲーリングのようにスマートでゲッペルスのように背が高く
ヒトラーのようなブロンドのゲルマン民族ですね、分かります。
ヒトラーのようなブロンドのゲルマン民族ですね、分かります。
美形というにはあまりにごっつくむさいリアルゲルマン人
178 :世界@名無史さん:2012/10/06(土) 23:23:22.57 0
179 :世界@名無史さん:2012/10/06(土) 23:30:28.38 O
ゲルマン=アーリア人=高尚な人種=日本人=東方アーリア人
180 :世界@名無史さん:2012/10/06(土) 23:32:47.43 0
ゲルマンと日本人を一緒にするのは冒涜です。
やめてください。
神と犬を一緒に語るのと一緒。
やめてください。
神と犬を一緒に語るのと一緒。
アメリカは対日・対中貿易赤字が慢性的で国内生産喚起に躍起になっているというが。
182 :世界@名無史さん:2012/10/06(土) 23:39:03.83 0
ドイツ=十字軍=テンプル騎士団=ロンメル機構化師団(ロマンありすぎる。まさにファンタジー)
日本=元寇で命からがら=猿吉、大陸でフルボッコ
話にならないよ
日本=元寇で命からがら=猿吉、大陸でフルボッコ
話にならないよ
日本人=東方アーリア人 は
ヒムラー国家長官の承認事項だ。
ドイツ=劣等スラヴにベルリンレイプ
日本=大陸でチンピラゴロツキに連戦連勝
日本=大陸でチンピラゴロツキに連戦連勝
容貌的にはイラン>>スラヴ>>>>>>>>ゲルマン
(17歳まで)
(17歳まで)
爬虫類はカメレオンなんてカラフルで目を奪う。
保護色という生存本能だな。
弱い種目の生きる知恵。
保護色という生存本能だな。
弱い種目の生きる知恵。
188 :世界@名無史さん:2012/10/06(土) 23:48:29.30 0
>>183
そんなの、劣等スラブ対策で、名誉白人に認定したまでだよ。
>>184
ドイツ=カール大帝欧州統一
日本=大陸の軍閥政権程度にも勝利ならず。 大東亜共栄圏どころか近海も危機一発
もう規模が違う・・・
そんなの、劣等スラブ対策で、名誉白人に認定したまでだよ。
>>184
ドイツ=カール大帝欧州統一
日本=大陸の軍閥政権程度にも勝利ならず。 大東亜共栄圏どころか近海も危機一発
もう規模が違う・・・
189 :世界@名無史さん:2012/10/06(土) 23:57:04.57 0
>>186
人種がすくなすぎってなに?ぱかなの?
ユーラシアは多種多様でも欧州権では黒人はそれなりに居住権獲得してるが、
劣等東アジアンはまったくゴキブリ扱いで居住権すら与えられてないでしょうに。
欧州での東アジアンの生存率の少なさワロスwww雑魚丸出しww
>>187
その弱いとされる保護色白人は、歴史上最強なんですが?
>>185
容姿
ゲルマン=ノルマン=トルキー>>イラン=東欧>>スラブ(スラブ上げはゴミ東洋人のみ)>>
まーでも難しいね。アングロサクソンは不細工といわれるが、ウィリアム王子の女めちゃきれいだし、
スコッティーとの混血だからまあまあだよ。
人種がすくなすぎってなに?ぱかなの?
ユーラシアは多種多様でも欧州権では黒人はそれなりに居住権獲得してるが、
劣等東アジアンはまったくゴキブリ扱いで居住権すら与えられてないでしょうに。
欧州での東アジアンの生存率の少なさワロスwww雑魚丸出しww
>>187
その弱いとされる保護色白人は、歴史上最強なんですが?
>>185
容姿
ゲルマン=ノルマン=トルキー>>イラン=東欧>>スラブ(スラブ上げはゴミ東洋人のみ)>>
まーでも難しいね。アングロサクソンは不細工といわれるが、ウィリアム王子の女めちゃきれいだし、
スコッティーとの混血だからまあまあだよ。
共産主義撲滅・東方生存権獲得どころか劣等民族にフルボッコされ共産主義拡大・国家分断に至った偉大なるアーリア人種
結論としては
白人動物的に美形(例:カメレオン)
能力はまあまあ
戦闘は弱い
総合点は10点中8点くらい
白人動物的に美形(例:カメレオン)
能力はまあまあ
戦闘は弱い
総合点は10点中8点くらい
193 :世界@名無史さん:2012/10/07(日) 00:08:14.88 0
>>190
それは違う!!
ドイツは、その前に電撃作戦により攻略付加と歌われたフランスマジノ要塞を3ヶ月で攻略。
さらに、イギリスを世界で初にかいはつした大陸間弾道弾で焦土作戦で降伏寸前に。
東欧諸国をまとめて数ヶ月で降伏に。
アフリカ戦線で連合軍に圧勝。
その上でのロシア進出。しかし、モスクワ100キロ手前のスターリングラードまで占領。
ロシア降伏寸前。
それに比べ日本は、ノモンハンで精鋭陸軍がたった一部隊に一日で壊滅・・・
本場アーリア人は名誉アーリア人とはわけが違う。
>>191
大和民族も渡来(逃げ)弥生人国家でしょうにwww
それは違う!!
ドイツは、その前に電撃作戦により攻略付加と歌われたフランスマジノ要塞を3ヶ月で攻略。
さらに、イギリスを世界で初にかいはつした大陸間弾道弾で焦土作戦で降伏寸前に。
東欧諸国をまとめて数ヶ月で降伏に。
アフリカ戦線で連合軍に圧勝。
その上でのロシア進出。しかし、モスクワ100キロ手前のスターリングラードまで占領。
ロシア降伏寸前。
それに比べ日本は、ノモンハンで精鋭陸軍がたった一部隊に一日で壊滅・・・
本場アーリア人は名誉アーリア人とはわけが違う。
>>191
大和民族も渡来(逃げ)弥生人国家でしょうにwww
欧州人
体臭:きつい
体毛:濃い
骨格:ごつい・年をとると肥える
体臭:きつい
体毛:濃い
骨格:ごつい・年をとると肥える
>モスクワ100キロ手前のスターリングラードまで占領。
どこの世界の話をしてるんだ・・・パラレルワールドって奴?
どこの世界の話をしてるんだ・・・パラレルワールドって奴?
なんでそんなに逃げ足が速いんだ白人
198 :世界@名無史さん:2012/10/07(日) 00:20:56.45 0
>>195
ゆとりパカは勉強しなさい!
>>196
あの宗教は大陸続きの世界最強激戦区の国々が密集する地域で、
政教一致による国家形成がなければ、過去フランク王国や神聖ローマ帝国の建国などありえなかった。
要はヒトラーもそうだが、人身掌握にたけた、まさに天才集団。
本当に感心する。
にくらべ、正月、クリスマス、パンプキン、仏教 神道、なんでも白人様のものはかっこいいとぱくる、
なにひとつ芯が通らない東洋のアーリア人とか寝言こいてる東洋民族は知ってる。
ゆとりパカは勉強しなさい!
>>196
あの宗教は大陸続きの世界最強激戦区の国々が密集する地域で、
政教一致による国家形成がなければ、過去フランク王国や神聖ローマ帝国の建国などありえなかった。
要はヒトラーもそうだが、人身掌握にたけた、まさに天才集団。
本当に感心する。
にくらべ、正月、クリスマス、パンプキン、仏教 神道、なんでも白人様のものはかっこいいとぱくる、
なにひとつ芯が通らない東洋のアーリア人とか寝言こいてる東洋民族は知ってる。
>ゆとりパカは勉強しなさい!
自己紹介はいいから
自己紹介はいいから
200 :世界@名無史さん:2012/10/07(日) 00:28:18.02 0
東洋人
口臭:歯並び最悪の口臭民族
体格:非常に劣る。ちび。
全体的:奇形 見劣りする。
口臭:歯並び最悪の口臭民族
体格:非常に劣る。ちび。
全体的:奇形 見劣りする。
黄色人種の日本人的のっぺり顔が広く王侯貴族階級として浸透して来たユーラシアでは、
のっぺり顔に対する畏怖が白人の遺伝子本能に埋め込まれている。
おそらく騎乗技術の発祥から数千年来の関係。
ユーラシア貴族の顔
http://www.kanteiya.com/famousepeople/images/Lenin/Lenin.jpg
ジョチ家の子孫(左)
http://upload.wikimedia.org/wikipedia/kk/8/82/Valikhanov.jpg
のっぺり顔に対する畏怖が白人の遺伝子本能に埋め込まれている。
おそらく騎乗技術の発祥から数千年来の関係。
ユーラシア貴族の顔
http://www.kanteiya.com/famousepeople/images/Lenin/Lenin.jpg
{kind=link}
ジョチ家の子孫(左)
http://upload.wikimedia.org/wikipedia/kk/8/82/Valikhanov.jpg
{kind=link}
202 :世界@名無史さん:2012/10/07(日) 00:34:29.06 0
日本も、捨てたもんじゃないと思うのは、日英同盟。
あれほど人類最強国家に並び証され、プライド最強人種たる、御イギリス様に、
東洋人の分際で不平等とはいえ同盟を結んでいただけたのは、日本至上最高の誇り!!!
本当に御エゲレス様には感謝感激と思うべき。
あれほど人類最強国家に並び証され、プライド最強人種たる、御イギリス様に、
東洋人の分際で不平等とはいえ同盟を結んでいただけたのは、日本至上最高の誇り!!!
本当に御エゲレス様には感謝感激と思うべき。
ウホウホやていたゲルマン人はユダヤ人のキリスト教によって人間らしく文明化された。
伍長閣下はこの事実を認めたくなくて消し去ろうと決意した。ユダヤ人など存在しない、と。
伍長閣下はこの事実を認めたくなくて消し去ろうと決意した。ユダヤ人など存在しない、と。
帝政ロシアのユダヤ人迫害に対抗するためユダヤ資本は日本へ資金を貸し付けた。
しかしナチのユダヤ人迫害には英米のユダヤ資本はほぼ傍観していた。
しかしナチのユダヤ人迫害には英米のユダヤ資本はほぼ傍観していた。
こんにちわ
私がイエス・キリストです。
http://28.media.tumblr.com/tumblr_lr8epxithG1qzo3c9o1_400.jpg
白人弟子のみなさん、変わった顔してますね。
私がイエス・キリストです。
http://28.media.tumblr.com/tumblr_lr8epxithG1qzo3c9o1_400.jpg
{kind=link}
白人弟子のみなさん、変わった顔してますね。
207 :世界@名無史さん:2012/10/07(日) 00:42:04.50 0
>>201
ユーラシアといわず、スラブのみにしてくれないか?
のっぺり不細工に遺伝的恐怖心抱いてるのはスラブ程度。
>>203
それはどうかな。ローマ大帝国を滅ぼしたものこそ神に導かれし民族げるまん
ユダヤは単にベルサイユ条約貴下のドイツ圧迫政治の鬱憤をユダヤ追討に向け、人心を掌握したかったまで。
ドイツ第三帝国構想範囲みれば、いかにゲルマン盟主の神聖ローマ帝国が、
偉大かつ盛大だったかわかる。もはやユダヤの亡霊など、中世ゲルマン史をみれば、
そんな劣等感など存在すべきはずがない。
ユーラシアといわず、スラブのみにしてくれないか?
のっぺり不細工に遺伝的恐怖心抱いてるのはスラブ程度。
>>203
それはどうかな。ローマ大帝国を滅ぼしたものこそ神に導かれし民族げるまん
ユダヤは単にベルサイユ条約貴下のドイツ圧迫政治の鬱憤をユダヤ追討に向け、人心を掌握したかったまで。
ドイツ第三帝国構想範囲みれば、いかにゲルマン盟主の神聖ローマ帝国が、
偉大かつ盛大だったかわかる。もはやユダヤの亡霊など、中世ゲルマン史をみれば、
そんな劣等感など存在すべきはずがない。
ヒャッハー・フン族にどつきまくられたウホウホ・ゲルマン人は
ユダヤの神に救いを求めローマに救いを求めて流民となってローマ領内に逃げ込んだ。
ユダヤの神に救いを求めローマに救いを求めて流民となってローマ領内に逃げ込んだ。
210 :世界@名無史さん:2012/10/07(日) 00:50:08.47 0
>>204
有色人種といってもモンゴロイドじゃないでしょwwww
コーカソイド有色人種ではあっても、モンゴロイドの分際が、でしゃばって言う資格なし。www
要はのっぺり不細工民族の言えた義理じゃないということだwww
>>205
そこにいたるほど、ドイツをベルサイユ条約などで徹底的に圧迫したのはそれら欧米列強です。
要はそれほど御ドイツ様が怖くて仕方なかったということ。
そのとおり。一次対戦から数年でまたもや、全世界相手に一カ国であそこまで戦えるほどの科学技術力を作り上げてしまう国。
ドイツ周辺の列強国が、ドイツをそうさせたんです。
有色人種といってもモンゴロイドじゃないでしょwwww
コーカソイド有色人種ではあっても、モンゴロイドの分際が、でしゃばって言う資格なし。www
要はのっぺり不細工民族の言えた義理じゃないということだwww
>>205
そこにいたるほど、ドイツをベルサイユ条約などで徹底的に圧迫したのはそれら欧米列強です。
要はそれほど御ドイツ様が怖くて仕方なかったということ。
そのとおり。一次対戦から数年でまたもや、全世界相手に一カ国であそこまで戦えるほどの科学技術力を作り上げてしまう国。
ドイツ周辺の列強国が、ドイツをそうさせたんです。
ノッペリ顔のヒムラー長官がチベット仏教に傾倒した、までは分かった
{kind=link}
213 :世界@名無史さん:2012/10/07(日) 00:57:50.76 0
>>208
ゆとりパカはもっと勉強に励め!!
>>209
当時のフン族の強さは異常!!
あんなのの最強ハンムラビだって勝てないよ。
フン族に負けたのは恥じらいじゃない。
近代西洋列強白人様にどつきたおされた東洋土人のほうが、よほど情けないw。
わずか黒船数席に屈した土人も情けないわ・・・w
だが、東洋の土民とはまったく無関係。
ゆとりパカはもっと勉強に励め!!
>>209
当時のフン族の強さは異常!!
あんなのの最強ハンムラビだって勝てないよ。
フン族に負けたのは恥じらいじゃない。
近代西洋列強白人様にどつきたおされた東洋土人のほうが、よほど情けないw。
わずか黒船数席に屈した土人も情けないわ・・・w
だが、東洋の土民とはまったく無関係。
ウホウホゲルマン人の分際が
我々アーリア人が人類の全ての文化・文明を創造した!キリ
なんて言うほうが出すぎたまねだと
我々アーリア人が人類の全ての文化・文明を創造した!キリ
なんて言うほうが出すぎたまねだと
>>213
白色民族は
有色人種の教えを信じ、
モンゴロイドに畏怖を覚え、
イスラムから逃げて海を渡り、
今日まで生きて来たのだ。弱小品種の生きる苦労がうかがいしれる。
見た目はよくても中身は空っぽ。
白色民族は
有色人種の教えを信じ、
モンゴロイドに畏怖を覚え、
イスラムから逃げて海を渡り、
今日まで生きて来たのだ。弱小品種の生きる苦労がうかがいしれる。
見た目はよくても中身は空っぽ。
白人に同情票を一票
217 :世界@名無史さん:2012/10/07(日) 01:04:35.22 0
>>212
全然違うが、
まーかりに日本人で何百万人に一人の顔をもってきて、モンゴロイドと代わらないって言われてもなw
のっぺり土人国家であの顔も何百万人に一人だが?w
それに、君が言ってるのは、なんだっけ?ツイッターかなんかの開発者のアメリカ白人様と、
お笑い芸人のサバンナ高橋が似てるって話題になったけど、そのレベルの話だよw
ノッチとオバマがにてる!といってるレベルよwイエロー民族哀れw
>>211
のっぺり不細工民族が、対ロシアのなんか作戦にびびりたおし、
人類最強人種に泣きつて同盟結んでもらったまで読んだ。
全然違うが、
まーかりに日本人で何百万人に一人の顔をもってきて、モンゴロイドと代わらないって言われてもなw
のっぺり土人国家であの顔も何百万人に一人だが?w
それに、君が言ってるのは、なんだっけ?ツイッターかなんかの開発者のアメリカ白人様と、
お笑い芸人のサバンナ高橋が似てるって話題になったけど、そのレベルの話だよw
ノッチとオバマがにてる!といってるレベルよwイエロー民族哀れw
>>211
のっぺり不細工民族が、対ロシアのなんか作戦にびびりたおし、
人類最強人種に泣きつて同盟結んでもらったまで読んだ。
218 :世界@名無史さん:2012/10/07(日) 01:13:21.64 0
>>214
問題はアーリア人が期限かじゃない。
古代アーリア人をしのぐ偉業を成し遂げた中世、そして近代ゲルマン民族が、
自らを、アーリア人の末裔、神に選ばれし民族神授説を唱えたとしてもなんらおかしいことじゃない。
>.215
極東の島に引きこもってきたイエローモンキーの末裔どもが言えた義理じゃないなwwww
西洋中東は陸続きの、人類際激戦区ですぞ?
時に逃げ、時に制覇し、そして、もはや銀河系最強生物たるゲルマン民族の歴史がある。
東洋の引きこもりは、あの激戦区で勝ち残る壮絶さはわからんだろうな〜黄猿どもwww
黒船3席に開国してる場合じゃありませんぞwwwwムッフォッフォッフォwwwイエローモンクゥィ〜ww
問題はアーリア人が期限かじゃない。
古代アーリア人をしのぐ偉業を成し遂げた中世、そして近代ゲルマン民族が、
自らを、アーリア人の末裔、神に選ばれし民族神授説を唱えたとしてもなんらおかしいことじゃない。
>.215
極東の島に引きこもってきたイエローモンキーの末裔どもが言えた義理じゃないなwwww
西洋中東は陸続きの、人類際激戦区ですぞ?
時に逃げ、時に制覇し、そして、もはや銀河系最強生物たるゲルマン民族の歴史がある。
東洋の引きこもりは、あの激戦区で勝ち残る壮絶さはわからんだろうな〜黄猿どもwww
黒船3席に開国してる場合じゃありませんぞwwwwムッフォッフォッフォwwwイエローモンクゥィ〜ww
ナチ時代ゲルマン人はユダヤ人の宗教であるキリスト教の戒律を嫌い
SS隊員の6割以上がキリスト教を棄教していた。
やっぱウホウホして十字きってるより東部戦線でヒャッハーしたいもんね。
SS隊員の6割以上がキリスト教を棄教していた。
やっぱウホウホして十字きってるより東部戦線でヒャッハーしたいもんね。
>古代アーリア人をしのぐ偉業を成し遂げた中世、そして近代ゲルマン民族が、
>自らを、アーリア人の末裔、神に選ばれし民族神授説を唱えたとしてもなんらおかしいことじゃない
で東方劣等民族を根絶しようとして返り討ちにされ
フルボッコされてベルリンレイプ、国家分断と。
>自らを、アーリア人の末裔、神に選ばれし民族神授説を唱えたとしてもなんらおかしいことじゃない
で東方劣等民族を根絶しようとして返り討ちにされ
フルボッコされてベルリンレイプ、国家分断と。
223 :世界@名無史さん:2012/10/07(日) 01:22:39.96 0
>>219
容姿はもうねwwww
いいかい?チンパンジーが人間に、「あいつ不細工じゃね?}って言ってたらどう思う?ww
珍パンはまだ動物園の人気者だが、白人様どころか、ネグロイドも、新モンゴロイドの突然変異ぶりは、
笑えないレベルだと思ってらっしゃるんですぞ?ww
同じ不細工でも、猿はかわいいし、笑える。ジョークにできる。
東洋黄色猿は笑えないからwwwリアルすぎる不細工民族www
>.220
まープロテスタント発祥の国だからね。キリスト教を破棄したのではなく、
組織を否定したまで。
ゲルマン様は懸命だよ。
なにやってもすごいよ。歴史刻んじゃうんだもん
容姿はもうねwwww
いいかい?チンパンジーが人間に、「あいつ不細工じゃね?}って言ってたらどう思う?ww
珍パンはまだ動物園の人気者だが、白人様どころか、ネグロイドも、新モンゴロイドの突然変異ぶりは、
笑えないレベルだと思ってらっしゃるんですぞ?ww
同じ不細工でも、猿はかわいいし、笑える。ジョークにできる。
東洋黄色猿は笑えないからwwwリアルすぎる不細工民族www
>.220
まープロテスタント発祥の国だからね。キリスト教を破棄したのではなく、
組織を否定したまで。
ゲルマン様は懸命だよ。
なにやってもすごいよ。歴史刻んじゃうんだもん
>>223
遊牧民が馬に乗って突然やって来るのと同じで、
南蛮人が船に乗って突然やってきたときは
常識人日本人もずいぶん驚いた。
平和に暮らす文明人としては野蛮人には対抗する術は無い。
結局彼らは自分が発明したわけでもない蒸気機関車の模型やらなにやら置いていった。
こっちは全て自分の実力、向こうは他のメンバーの実績を自分のものとして紹介。
狡猾な野蛮人には手も足も出ない。
遊牧民が馬に乗って突然やって来るのと同じで、
南蛮人が船に乗って突然やってきたときは
常識人日本人もずいぶん驚いた。
平和に暮らす文明人としては野蛮人には対抗する術は無い。
結局彼らは自分が発明したわけでもない蒸気機関車の模型やらなにやら置いていった。
こっちは全て自分の実力、向こうは他のメンバーの実績を自分のものとして紹介。
狡猾な野蛮人には手も足も出ない。
古代、ウホウホ・ゲルマン人はフン族にヒャッハーとフルボッコされたが
ユダヤ人の教えに救いを求め、ローマ帝国の足元にすがって
ローマのインフラに寄生してようやく食いつなぐことが出来た。
近代においてゲルマン人は厚かましくもユダヤの教えに背き
ローマの恩人をパスタ野郎呼ばわりし、血迷ったのか東方フン族を
根絶したるわ劣等民族めと絶滅戦争を挑んだ。
ユダヤ人の教えに救いを求め、ローマ帝国の足元にすがって
ローマのインフラに寄生してようやく食いつなぐことが出来た。
近代においてゲルマン人は厚かましくもユダヤの教えに背き
ローマの恩人をパスタ野郎呼ばわりし、血迷ったのか東方フン族を
根絶したるわ劣等民族めと絶滅戦争を挑んだ。
しょうがないので白人は容姿を売って生きていけ。
キャバクラ嬢みたいに。
キャバクラ嬢みたいに。
*匂いきついです
228 :世界@名無史さん:2012/10/07(日) 01:30:13.74 0
>>221
気味ね、一次対戦、二次大戦と、ほぼ一カ国で全欧州、全世界を相手に、
あれだけ暴れまわれる国ほかにありますか?
あれ、どこかの東洋のアーリア人さんは、ホームグランドでアメリカ艦隊にフルボッコじゃなかったでしたっけ?ww
当方劣等民族といってもコーカソイドでしょw
モンゴロイドの劣等主に負けたわけじゃない。
しかも負けた原因は「冬将軍」!!!!!!!!!勘違いしちゃいかん!!
神風じゃもう、軍艦は重たいから沈まないよ?きづいてちょんまげwww
>>222
すらぶなんぞ、全然だめだめ。ただ単に気候と地理を利用しての偶発的勝利だらけ。
ひとつも実力なんて伴ってない。
それに比べ、神はバリバリじゃん?
めっちゃ全欧州を従え、オスマンとゲームの世界のような遠征繰り広げてるじゃん。
もう、戦っても絵になるっていうの?もう芸術の世界。
なにをやっても、芸術の域に達するのが神ゲルマン民族。
気味ね、一次対戦、二次大戦と、ほぼ一カ国で全欧州、全世界を相手に、
あれだけ暴れまわれる国ほかにありますか?
あれ、どこかの東洋のアーリア人さんは、ホームグランドでアメリカ艦隊にフルボッコじゃなかったでしたっけ?ww
当方劣等民族といってもコーカソイドでしょw
モンゴロイドの劣等主に負けたわけじゃない。
しかも負けた原因は「冬将軍」!!!!!!!!!勘違いしちゃいかん!!
神風じゃもう、軍艦は重たいから沈まないよ?きづいてちょんまげwww
>>222
すらぶなんぞ、全然だめだめ。ただ単に気候と地理を利用しての偶発的勝利だらけ。
ひとつも実力なんて伴ってない。
それに比べ、神はバリバリじゃん?
めっちゃ全欧州を従え、オスマンとゲームの世界のような遠征繰り広げてるじゃん。
もう、戦っても絵になるっていうの?もう芸術の世界。
なにをやっても、芸術の域に達するのが神ゲルマン民族。
そっくりやんけ
キャバ嬢
http://www.anzenadult.info/wp-content/uploads/2009/12/091216_sae.jpg
白人(微注意)
http://galleries.asstraffic.com/betty/03/s1/_content/16.jpg
キャバ嬢
http://www.anzenadult.info/wp-content/uploads/2009/12/091216_sae.jpg
{kind=link}
白人(微注意)
http://galleries.asstraffic.com/betty/03/s1/_content/16.jpg
{kind=link}
>気味ね、一次対戦、二次大戦と、ほぼ一カ国で全欧州、全世界を相手に、
>あれだけ暴れまわれる国ほかにありますか?
だから偉大なフン王アッティラの手下になって暴れてた話は知ってるって
>あれだけ暴れまわれる国ほかにありますか?
だから偉大なフン王アッティラの手下になって暴れてた話は知ってるって
232 :世界@名無史さん:2012/10/07(日) 01:39:12.30 0
>>224
常識人だったとは思えませんね〜〜
確かおバカな藩が総力挙げて海峡で砲撃した報復に、数日で砲台ごとフルボッコにされてましたね?www
常識人なら、自らやったんなら勝ちなさいよwww
エゲレス大使館を焼き討ちしたり生麦事件など、国際常識すら皆無だったというのが浮き彫りですよw
は・な・し・あ・いすら当時のご先祖さんたちしらなかったの?wwプププww
>>225
イエローもんきーの妄想乙ww
>>226
もうとっくにハリウッド映画という世界共通の文化で大活躍ですやんwww
白人様は何をやってもこうなんだよね〜〜〜wすごいと思わない?
猿的に早くあんな人間になりたいとか思わない?ww
>>227
東洋人も口臭きついですw
常識人だったとは思えませんね〜〜
確かおバカな藩が総力挙げて海峡で砲撃した報復に、数日で砲台ごとフルボッコにされてましたね?www
常識人なら、自らやったんなら勝ちなさいよwww
エゲレス大使館を焼き討ちしたり生麦事件など、国際常識すら皆無だったというのが浮き彫りですよw
は・な・し・あ・いすら当時のご先祖さんたちしらなかったの?wwプププww
>>225
イエローもんきーの妄想乙ww
>>226
もうとっくにハリウッド映画という世界共通の文化で大活躍ですやんwww
白人様は何をやってもこうなんだよね〜〜〜wすごいと思わない?
猿的に早くあんな人間になりたいとか思わない?ww
>>227
東洋人も口臭きついですw
納豆を国防軍の携帯食にしようと試みた伍長閣下。
>ハリウッド映画という世界共通の文化
その割には海外興行は赤字が多いな
その割には海外興行は赤字が多いな
235 :世界@名無史さん:2012/10/07(日) 01:49:39.27 0
>>206の画像ってなに?最新の研究で出たキリストの復元顔とか?
236 :世界@名無史さん:2012/10/07(日) 01:51:22.14 0
>>229
目まで劣化してるのイエローってwww
学説発表したら?ww
そもそもドンだけ画像処理してるのよそれww
>>231
自国の文化を否定してまで、脱亜入欧と発狂した国家なら知ってるってw
>>233
ほら!!みてみな!!脱亜入欧の典型例ww
自国の文化を否定する修正は直りませんな?ww
納豆なんて食べちゃう野蛮行為が許せないと言っちゃうお茶目さん♪
白人コンプ土人乙wwwwwww
目まで劣化してるのイエローってwww
学説発表したら?ww
そもそもドンだけ画像処理してるのよそれww
>>231
自国の文化を否定してまで、脱亜入欧と発狂した国家なら知ってるってw
>>233
ほら!!みてみな!!脱亜入欧の典型例ww
自国の文化を否定する修正は直りませんな?ww
納豆なんて食べちゃう野蛮行為が許せないと言っちゃうお茶目さん♪
白人コンプ土人乙wwwwwww
キリストの復元顔らしい
240 :世界@名無史さん:2012/10/07(日) 02:00:13.37 0
>>237
いやいやいや、お隣の大陸半島諸国と近親の島国渡来人国家にいわれましてもね〜〜ww♪
いいでしょう。仮にですよ?仮に白人が強引だと仮定しましょう。
で、その強引さの向こう側には文明開化があるとおっしゃったのも日本でそ?w
よくごらんなさい?その強引さと引き換えに、みな、東洋諸国は近代かいたしました。
それだけじゃない、御白人様様は、あらゆる額もにゃか嶽技術の産物、そして政治のあり方まで、
いえ、もうあらゆるものを与えてくださりました。
それに引き換え東洋のど人どもはどうでしょうww
モンゴルが過去スラブを占領しましたが、著しく文明は退化し、
西洋諸国から劣等と認識されるまでになってしまったスラブ。
東洋土人は、荒らしても何もその後にもたらしませんwww害悪そのものwww
同じ強引でもまったく副産物は違いますwww黄猿涙目wwwwww
いやいやいや、お隣の大陸半島諸国と近親の島国渡来人国家にいわれましてもね〜〜ww♪
いいでしょう。仮にですよ?仮に白人が強引だと仮定しましょう。
で、その強引さの向こう側には文明開化があるとおっしゃったのも日本でそ?w
よくごらんなさい?その強引さと引き換えに、みな、東洋諸国は近代かいたしました。
それだけじゃない、御白人様様は、あらゆる額もにゃか嶽技術の産物、そして政治のあり方まで、
いえ、もうあらゆるものを与えてくださりました。
それに引き換え東洋のど人どもはどうでしょうww
モンゴルが過去スラブを占領しましたが、著しく文明は退化し、
西洋諸国から劣等と認識されるまでになってしまったスラブ。
東洋土人は、荒らしても何もその後にもたらしませんwww害悪そのものwww
同じ強引でもまったく副産物は違いますwww黄猿涙目wwwwww
17歳まで限定ならどう見てもゲルマンよりスラヴ
そもそもモンゴルに関係なく西欧世界は東方正教を異端視していた
243 :世界@名無史さん:2012/10/07(日) 02:06:17.06 0
>>239
仕組みは違いますが似たようなもんですよ。w
ただ白人様はやることがダイナミックというだけw。
東洋人と白人がやると、ここまで規模が違う=世界の注目度合いの差
なんですね。白人様には全世界が興味心身で釘付け!という裏返しなんですね〜〜♪
仕組みは違いますが似たようなもんですよ。w
ただ白人様はやることがダイナミックというだけw。
東洋人と白人がやると、ここまで規模が違う=世界の注目度合いの差
なんですね。白人様には全世界が興味心身で釘付け!という裏返しなんですね〜〜♪
>御白人様様は、あらゆる額もにゃか嶽技術の産物、そして政治のあり方まで、
>いえ、もうあらゆるものを与えてくださりました
ユダヤの教えがウホウホ・ヨーロッパ人に人の道を教えた、までは読んだ
>いえ、もうあらゆるものを与えてくださりました
ユダヤの教えがウホウホ・ヨーロッパ人に人の道を教えた、までは読んだ
>>240
モンゴルは東西交通を開いたな。
そして被支配者に対して圧倒的に寛大。信仰は自由。文化には敬意。
ところが白人樣はその侵略方法の野蛮な部分のみを見習ったわけだ。
奴隷制もイスラムのパクり。一方はマムルークとして栄達の道があり
一方は陰惨な人権剥奪の歴史。
同じ制度で実施する人間が違うとこうも変わるものだろうか。
モンゴルは東西交通を開いたな。
そして被支配者に対して圧倒的に寛大。信仰は自由。文化には敬意。
ところが白人樣はその侵略方法の野蛮な部分のみを見習ったわけだ。
奴隷制もイスラムのパクり。一方はマムルークとして栄達の道があり
一方は陰惨な人権剥奪の歴史。
同じ制度で実施する人間が違うとこうも変わるものだろうか。
247 :世界@名無史さん:2012/10/07(日) 02:12:56.35 0
>>241
それはモンゴロイドゆえのサガですねwww
スラブなんて西洋では奇形扱い。
のっぺりしてるからです。w
>.242
まーそれも一因としてはあったでしょうがね。
どうも、東洋と接する地域は、非常に文明が劣化するというのはもう証明できますよねw
過去の中東も、あれだけの文化を誇りながら、西洋と比べ、劣化率は凄まじい。
東洋、東方とかかわるとろくなことにならないという見本ですよねwww。
西欧列強さま様も、近代、東洋に進出してからが運のつき。経済から文化まで廃れ始めていった。
東洋というのは、百害あって一理なしという典型ですねww
私は白人様に東洋の地には、けがれるからふみいれないでほちい。
白人様の血を守りたい。
それはモンゴロイドゆえのサガですねwww
スラブなんて西洋では奇形扱い。
のっぺりしてるからです。w
>.242
まーそれも一因としてはあったでしょうがね。
どうも、東洋と接する地域は、非常に文明が劣化するというのはもう証明できますよねw
過去の中東も、あれだけの文化を誇りながら、西洋と比べ、劣化率は凄まじい。
東洋、東方とかかわるとろくなことにならないという見本ですよねwww。
西欧列強さま様も、近代、東洋に進出してからが運のつき。経済から文化まで廃れ始めていった。
東洋というのは、百害あって一理なしという典型ですねww
私は白人様に東洋の地には、けがれるからふみいれないでほちい。
白人様の血を守りたい。
鑑賞用ペットとして品種保持せねばならんよな。
>私は白人様に東洋の地には、けがれるからふみいれないでほちい。
新大陸やオセアニアの入植者の悪口はそこまでだ。
新大陸やオセアニアの入植者の悪口はそこまでだ。
250 :世界@名無史さん:2012/10/07(日) 02:25:07.68 0
>>244
仮に層だとしても、アインシュタインはドイツ生まれには変わりありませんからwwww
ユダヤって日本に居たの?wwあれれ?消えた十氏族妄想のユダヤ人はいるの???プププwww
>>245
それは、東洋人は宗教による価値観は、信仰心が希薄で信心深くないからです。
白人様は、非常に信仰心が厚く、道徳心があるからの違いです。
東洋人のように信仰心がないから、日本は逆に脱亜入欧など、他社文化を受け入れやすかったんでしょう。
要は他社に寛容=信仰心で抑制することがない残虐非道な侵略者でもあったから、あれだけ広大な地域支配できたのでしょう。
人権略奪ですが、世界で人権宣言出した国は御フランス様ですよ?www
国王や皇帝は法により支配されると定めてくれた文明人も白人様ですよ?www
あなたらイエローもん黄〜〜どもは、近代までなにをやってきたの?wwww
白人様が新しい近代ルール全部、血と汗と涙のすえに勝ち取ってくださったんでしょwww
まったく、東洋の未開猿は、何一つ人類の歴史に貢献してませんなwwww土人猿乙www
>。246
世界的シェアでのネッッとエロ画像も白人様でしょうにww
東洋人なんて、外人の世界ではいわゆるキワモノ扱いでしょうにwww
仮に層だとしても、アインシュタインはドイツ生まれには変わりありませんからwwww
ユダヤって日本に居たの?wwあれれ?消えた十氏族妄想のユダヤ人はいるの???プププwww
>>245
それは、東洋人は宗教による価値観は、信仰心が希薄で信心深くないからです。
白人様は、非常に信仰心が厚く、道徳心があるからの違いです。
東洋人のように信仰心がないから、日本は逆に脱亜入欧など、他社文化を受け入れやすかったんでしょう。
要は他社に寛容=信仰心で抑制することがない残虐非道な侵略者でもあったから、あれだけ広大な地域支配できたのでしょう。
人権略奪ですが、世界で人権宣言出した国は御フランス様ですよ?www
国王や皇帝は法により支配されると定めてくれた文明人も白人様ですよ?www
あなたらイエローもん黄〜〜どもは、近代までなにをやってきたの?wwww
白人様が新しい近代ルール全部、血と汗と涙のすえに勝ち取ってくださったんでしょwww
まったく、東洋の未開猿は、何一つ人類の歴史に貢献してませんなwwww土人猿乙www
>。246
世界的シェアでのネッッとエロ画像も白人様でしょうにww
東洋人なんて、外人の世界ではいわゆるキワモノ扱いでしょうにwww
品評会
白人
http://upload.wikimedia.org/wikipedia/commons/thumb/8/81/Europaeid_types.jpg/640px-Europaeid_types.jpg
ペルシャ猫
http://nekonoyume.com/gazou/syurui/40.gif
白人
http://upload.wikimedia.org/wikipedia/commons/thumb/8/81/Europaeid_types.jpg/640px-Europaeid_types.jpg
{kind=link}
ペルシャ猫
http://nekonoyume.com/gazou/syurui/40.gif
{kind=link}
>人権略奪ですが、世界で人権宣言出した国は御フランス様ですよ?www
>国王や皇帝は法により支配されると定めてくれた文明人も白人様ですよ?
なんのロベスピエールの話だ?
>国王や皇帝は法により支配されると定めてくれた文明人も白人様ですよ?
なんのロベスピエールの話だ?
エロ画像の需要を自慢しとるわコイツ。
AV女優=白人
「白人」で画像ググるとエロ画像しか出て来なかったわ。
「日本人」でググると書籍とか著名人が出て来た。
AV女優=白人
「白人」で画像ググるとエロ画像しか出て来なかったわ。
「日本人」でググると書籍とか著名人が出て来た。
>アインシュタインはドイツ生まれには変わりありませんから
パレスチナ生まれのユダヤ人大工の息子の教えなんだが
パレスチナ生まれのユダヤ人大工の息子の教えなんだが
255 :世界@名無史さん:2012/10/07(日) 02:32:13.23 0
いや〜〜〜〜久々に完全論破 独走状態でしたねwwww
白人様にあらずんば、ひとにあらず!!
昔平家が言った言葉です。ww
みなさん、白人様に生まれられなかった時点で罰ゲームです。自覚しましょうね♪w
白人様にあらずんば、ひとにあらず!!
昔平家が言った言葉です。ww
みなさん、白人様に生まれられなかった時点で罰ゲームです。自覚しましょうね♪w
>>250
エロ白人は日本に略奪に来ただけではないか。
モンゴルは東洋人じゃねえぞ。
なぜ他者に寛容がマイナス要因にされてるのか理解に苦しむ。
白人理論では他民族を拷問とかやっちゃう方が信仰心に満ちてることになるようだ。
さすがカルト宗教キリスト教(有色)
エロ白人は日本に略奪に来ただけではないか。
モンゴルは東洋人じゃねえぞ。
なぜ他者に寛容がマイナス要因にされてるのか理解に苦しむ。
白人理論では他民族を拷問とかやっちゃう方が信仰心に満ちてることになるようだ。
さすがカルト宗教キリスト教(有色)
> 白人様にあらずんば、ひとにあらず!!
> 昔平家が言った言葉です。ww
これが白人代表とは・・・
たまげた馬鹿だ
> 昔平家が言った言葉です。ww
これが白人代表とは・・・
たまげた馬鹿だ
258 :世界@名無史さん:2012/10/07(日) 02:38:28.90 0
>>252
ごまかさないでくださいw
イギリスの偉大な歴史をごまかさないでくださいw。
そしてフランスの偉大な革命。
おそらく世界に東洋しか存在しなかったら、100億年たってもフランス革命のようなのはおきなかったでしょうねw。
猿の惑星のような中世世界ですよ100億年後もwww
>>253
図書館に行って学問書を手にとって見なさい。
あらゆるこの世の学問の期限は白人様。
もはやあいた口がふさがらないレベル。
いい?空を見上げ、綺麗だなと思い、宇宙の仕組みを研究し、ほんとに宇宙間で言っちゃうのが御白人様w
空を見上げ、綺麗だなと思い、月にはウサギがすんでると妄想して酒飲むのが東洋土人wwwwww
もう全然違うのwwwwwwwwwwwwwwwwww
ごまかさないでくださいw
イギリスの偉大な歴史をごまかさないでくださいw。
そしてフランスの偉大な革命。
おそらく世界に東洋しか存在しなかったら、100億年たってもフランス革命のようなのはおきなかったでしょうねw。
猿の惑星のような中世世界ですよ100億年後もwww
>>253
図書館に行って学問書を手にとって見なさい。
あらゆるこの世の学問の期限は白人様。
もはやあいた口がふさがらないレベル。
いい?空を見上げ、綺麗だなと思い、宇宙の仕組みを研究し、ほんとに宇宙間で言っちゃうのが御白人様w
空を見上げ、綺麗だなと思い、月にはウサギがすんでると妄想して酒飲むのが東洋土人wwwwww
もう全然違うのwwwwwwwwwwwwwwwwww
259 :世界@名無史さん:2012/10/07(日) 02:45:11.34 0
>>256
モンゴルも東洋人てくくりですよ白人様はw。
仮に中央アジアであっても、そういうくくりで判断してない余白人さまは。
顔立ちがのっぺり不細工ならみな東洋人wwwww例外なく東洋人wwwww
アジアンとは中東コーカソイドのことだからね。
あくまで、モンゴル人の宗教に寛容といういみは、他人種侵略なため、
寛容さが必要だっただけ。それだけですよ。
オスマン帝国も寛容だった。それはまったくの異民族異人種支配だったから。
西洋が、拷問といういめーじになるのは、近親である欧州権内の近親民族のしハイなため、
徳川のキリシタン弾圧のように恐怖心からのもの。
まったく意味合いが違いますねw
モンゴルも東洋人てくくりですよ白人様はw。
仮に中央アジアであっても、そういうくくりで判断してない余白人さまは。
顔立ちがのっぺり不細工ならみな東洋人wwwww例外なく東洋人wwwww
アジアンとは中東コーカソイドのことだからね。
あくまで、モンゴル人の宗教に寛容といういみは、他人種侵略なため、
寛容さが必要だっただけ。それだけですよ。
オスマン帝国も寛容だった。それはまったくの異民族異人種支配だったから。
西洋が、拷問といういめーじになるのは、近親である欧州権内の近親民族のしハイなため、
徳川のキリシタン弾圧のように恐怖心からのもの。
まったく意味合いが違いますねw
261 :世界@名無史さん:2012/10/07(日) 02:54:19.07 0
>>260
中二病でも、最後は実現さしちゃうのが白人様クオリティーなんですよねw
だってさ、月が綺麗、→つきってどうなってるの?→宗教的世界を否定しても仕組みを暴く(ガリレオ)→月の表面見てみたい→天体望遠鏡作る→いっちまおうぜ!!→ホントにロケットで到達。
こんな発想東洋土人にできますか?qwwww
東洋土人は、
月が綺麗→なんかウサギが見えるね→饅頭とススキあげて和歌詠もう→ 100万年後・・・今日もお月様綺麗だね♪さあ和歌詠もう♪www
中二病でも、最後は実現さしちゃうのが白人様クオリティーなんですよねw
だってさ、月が綺麗、→つきってどうなってるの?→宗教的世界を否定しても仕組みを暴く(ガリレオ)→月の表面見てみたい→天体望遠鏡作る→いっちまおうぜ!!→ホントにロケットで到達。
こんな発想東洋土人にできますか?qwwww
東洋土人は、
月が綺麗→なんかウサギが見えるね→饅頭とススキあげて和歌詠もう→ 100万年後・・・今日もお月様綺麗だね♪さあ和歌詠もう♪www
>>259
ウラル山脈かドニエプル川かいう意味を成さない境界のことか。
逃げる前にはゲルマンが住んでたあたりのことな。
>>261
やっぱ東洋人の方がマトモだ。
月まで逃げるなよ。恐がりにもほどがある。
ウラル山脈かドニエプル川かいう意味を成さない境界のことか。
逃げる前にはゲルマンが住んでたあたりのことな。
>>261
やっぱ東洋人の方がマトモだ。
月まで逃げるなよ。恐がりにもほどがある。
大体あれ月に本当に行ってないでしょ?
インチキとさんざん言われてるじゃん。
空気も無いのに旗がはためいたり
撮影の背景セットで違う地点で同じ石を使ってたり。
インチキとさんざん言われてるじゃん。
空気も無いのに旗がはためいたり
撮影の背景セットで違う地点で同じ石を使ってたり。
地球の周り上空数百キロのトコを無人で回ってるだけなのが、
いきなり電子計算機も無い時代に38万キロ飛び出すなんて理にかなってねえし。
数十年経った今も上空で数日暮らしただけの実験を大ニュースとして伝えるくらいなのに、
1968年に月面まで言ってるかあ?
いきなり電子計算機も無い時代に38万キロ飛び出すなんて理にかなってねえし。
数十年経った今も上空で数日暮らしただけの実験を大ニュースとして伝えるくらいなのに、
1968年に月面まで言ってるかあ?
265 :世界@名無史さん:2012/10/07(日) 03:08:15.21 0
>>262
それを逆に返せば、逃げたということではなく、東方に見切りをつけたと思いませんか?
ゲルマン人が東洋に居座ってたら、今の世界は存在しない。東洋の地に染まったのっぺり不細工劣等種wwwwwなんて創造したくないものwww
それは神のお導きなんですよ。歴史が適正化されたということ。
それから、よーーくかんがえてみなさい。www
中世のように現代でも白人様が民族粛清できるなら、東洋土人は根絶やしでしょうねwwww
しかし、それができない今となっては、歴史に何の貢献も影響も及ぼさず、
ただ東洋の地に引きこもってばかばかと人数だけ増やしてきたゴキブリ東洋人のおかげで、
もう地球の恵みはあと何年も持ちませんw
そのためにまた白人様は、自らの血と汗と努力の末に、新たに宇宙空間に人間のフロンティアを構想してくれてるんです。
誰のおかげだと思ってるの?wwwゴキブリ東洋人のおかげで宇宙開拓をしてくれてるんですwww
白人様、ほんと〜〜〜〜〜〜〜〜に東洋土人のおかげで、ご苦労をおかけします!!!w
白人様に栄光あれ!!
それを逆に返せば、逃げたということではなく、東方に見切りをつけたと思いませんか?
ゲルマン人が東洋に居座ってたら、今の世界は存在しない。東洋の地に染まったのっぺり不細工劣等種wwwwwなんて創造したくないものwww
それは神のお導きなんですよ。歴史が適正化されたということ。
それから、よーーくかんがえてみなさい。www
中世のように現代でも白人様が民族粛清できるなら、東洋土人は根絶やしでしょうねwwww
しかし、それができない今となっては、歴史に何の貢献も影響も及ぼさず、
ただ東洋の地に引きこもってばかばかと人数だけ増やしてきたゴキブリ東洋人のおかげで、
もう地球の恵みはあと何年も持ちませんw
そのためにまた白人様は、自らの血と汗と努力の末に、新たに宇宙空間に人間のフロンティアを構想してくれてるんです。
誰のおかげだと思ってるの?wwwゴキブリ東洋人のおかげで宇宙開拓をしてくれてるんですwww
白人様、ほんと〜〜〜〜〜〜〜〜に東洋土人のおかげで、ご苦労をおかけします!!!w
白人様に栄光あれ!!
266 :世界@名無史さん:2012/10/07(日) 03:14:21.62 0
>>263>>264
てれびに釣られすぎww
仮に当時がいけてなかったとしても、
もはや無人で火星言っちゃってる時代ですからねw。
つきなんてちょろいもんですよw
マー有人から無人観察というマクロ的価値転換でまた新たなマクロ築いちゃうのが白人様なんだなこれがww
馬鹿東洋人は、一回有人に成功したら、千年後も同じ手法でやってそうだからなwwwwwwwwww
てれびに釣られすぎww
仮に当時がいけてなかったとしても、
もはや無人で火星言っちゃってる時代ですからねw。
つきなんてちょろいもんですよw
マー有人から無人観察というマクロ的価値転換でまた新たなマクロ築いちゃうのが白人様なんだなこれがww
馬鹿東洋人は、一回有人に成功したら、千年後も同じ手法でやってそうだからなwwwwwwwwww
>>265
いや、スペイン人が大航海したのはオスマンの支配する地中海ルートを使いたくなかったからだし、
ヴェネチアやジェノヴァの航海術を利用したものだし、
それで開拓された航海ルートを使ったアングロサクソンなんてもはやパクり以外の何ものでもない。
探求心などかけらもないただの中央交易ルートからの逃亡。イスラム3帝国は強かったからね。
創造性ゼロ。
いや、スペイン人が大航海したのはオスマンの支配する地中海ルートを使いたくなかったからだし、
ヴェネチアやジェノヴァの航海術を利用したものだし、
それで開拓された航海ルートを使ったアングロサクソンなんてもはやパクり以外の何ものでもない。
探求心などかけらもないただの中央交易ルートからの逃亡。イスラム3帝国は強かったからね。
創造性ゼロ。
268 :世界@名無史さん:2012/10/07(日) 03:25:23.82 0
>>267
白人様と同列の中東コーカソイドも強いですからねw
モンゴロイドとちがい、コーカソイドは超優等民族ですよねwww
それを私は想像性がないとは思わないのですね?
たとえば日本も、近隣大陸半島と、経済が廃れてもかかわってきませんでした。
これ正解です。
欧州においては、あれだけの先進国が、ほんのわずかの四国や九州くらいの土地で、
何十カ国にも分断し、ひしめき合ってるんですよ?www
イスパニアは日本にもすごい影響をもたらしてますが、
あれだけちっさい国々が、危険なルートや、回避術としてのものは日本と同じ合理化です。
創造性がないとそれでもしたいなら、合理性といったものでしょうw
あ〜〜〜また世界史板を論破しまくりたくなってきたな〜〜〜wwww
白人様と同列の中東コーカソイドも強いですからねw
モンゴロイドとちがい、コーカソイドは超優等民族ですよねwww
それを私は想像性がないとは思わないのですね?
たとえば日本も、近隣大陸半島と、経済が廃れてもかかわってきませんでした。
これ正解です。
欧州においては、あれだけの先進国が、ほんのわずかの四国や九州くらいの土地で、
何十カ国にも分断し、ひしめき合ってるんですよ?www
イスパニアは日本にもすごい影響をもたらしてますが、
あれだけちっさい国々が、危険なルートや、回避術としてのものは日本と同じ合理化です。
創造性がないとそれでもしたいなら、合理性といったものでしょうw
あ〜〜〜また世界史板を論破しまくりたくなってきたな〜〜〜wwww
>>266
ソ連とやり合ってるってのがインチキくさいなあ。
あの国は捏造プロパガンダくらい平気でやりそうだろ。
それに対抗して捏造合戦になっててもおかしくなさそう。
じゃあなんでその時からもうやらないの?なんで?なんで?
ソ連とやり合ってるってのがインチキくさいなあ。
あの国は捏造プロパガンダくらい平気でやりそうだろ。
それに対抗して捏造合戦になっててもおかしくなさそう。
じゃあなんでその時からもうやらないの?なんで?なんで?
>>268
トルコ系にコーカソイドもモンゴロもあるかよ。
突厥はユーラシア全域民族。発祥は北東アジア。
イスパニア<オスマン<チムール<モンゴル
アングロサクソンはさらにそのまた蛮族だし。
逃げた先に偶然手つかずの資源があったという宝くじのおかげ。
トルコ系にコーカソイドもモンゴロもあるかよ。
突厥はユーラシア全域民族。発祥は北東アジア。
イスパニア<オスマン<チムール<モンゴル
アングロサクソンはさらにそのまた蛮族だし。
逃げた先に偶然手つかずの資源があったという宝くじのおかげ。
辺地のウホウホ・ゲルマンがパレスチナのユダヤのメシアにすがって
救いを求めてヨーロッパにキリスト教は根を下ろした
救いを求めてヨーロッパにキリスト教は根を下ろした
四国の土地に何十カ国はねえだろ。
この日本人ゲルマン馬鹿なのか。論破とか言い出す奴って低能しかいない。
この日本人ゲルマン馬鹿なのか。論破とか言い出す奴って低能しかいない。
「弥生人=おたふく・つり目」というのは間違い
それは朝鮮系です。
弥生人はモンゴロイドで、縄文人とほぼ似ています。これは最新の科学で明らかになっています。
それは朝鮮系です。
弥生人はモンゴロイドで、縄文人とほぼ似ています。これは最新の科学で明らかになっています。
つまり、
縄文人→弥生人→朝鮮人の3種があるということです日本人は。
縄文人→弥生人→朝鮮人の3種があるということです日本人は。
朝鮮系日本人は今の関西人(近畿人)です。
縄文弥生人は混血して沖縄と北海道と東北に生息します。アイヌなど。
なぜ南北に分かれたかというと朝鮮系日本人=大和朝廷に内地を追われたからです。(4世紀〜5世紀ごろ)
縄文弥生人は混血して沖縄と北海道と東北に生息します。アイヌなど。
なぜ南北に分かれたかというと朝鮮系日本人=大和朝廷に内地を追われたからです。(4世紀〜5世紀ごろ)
縄文人はオーストラロイドだよね?
278 :世界@名無史さん:2012/10/09(火) 10:06:30.63 0
ハプスブルク家は近親婚しまくってたのに何故美形揃いか。。そこに答えがある>ヨーロッパ人の容姿
>>273
弥生人も朝鮮人もY染色体O系統だから同類
弥生人も朝鮮人もY染色体O系統だから同類
280 :世界@名無史さん:2012/10/09(火) 17:17:34.62 0
スレ違いいいかげんにしろ
関西人は朝鮮人ってのは朝鮮では一般的な認識だよな。
同胞が日本を支配して誇らしいニダー
日本人のルーツは朝鮮人だよ
僕朝鮮人だよ
僕朝鮮人だよ
ウリナラ起源の大バーゲンも秋田
285 :世界@名無史さん:2012/10/17(水) 17:53:10.31 O
最近読んだ本によると
肌の浅黒いカンボジア人は比較的白いベトナム人を求め
肌の黒いケニア人は比較的ブラウンなエチオピア人を求めるらしい
あくまで買春の次元の話だけど
肌の浅黒いカンボジア人は比較的白いベトナム人を求め
肌の黒いケニア人は比較的ブラウンなエチオピア人を求めるらしい
あくまで買春の次元の話だけど
そりゃ、世界を制している人種は肌が白いからな。
オバマも実は結構白い白人だよね。
オバマも実は結構白い白人だよね。
白い黒人ねw
288 :世界@名無史さん:2012/10/18(木) 02:34:46.06 0
オバマが白かったら黒人に見えないよ
日本人もえたやひにんを虐殺すれば良かったのに
日本じゃえたひにんの方がきれいだよ。
芸能の職に就く者が多いし、隆慶一郎の世界のように異郷(西域人)の流浪民の末裔との発想で
碧眼や目鼻立ちがきれいという見方もある。
芸能の職に就く者が多いし、隆慶一郎の世界のように異郷(西域人)の流浪民の末裔との発想で
碧眼や目鼻立ちがきれいという見方もある。
野中広務
松本龍
山城新伍
どこが縄文系なんだ?
松本龍
山城新伍
どこが縄文系なんだ?
>>293
こいつら朝鮮系だろ
こいつら朝鮮系だろ
>>291
なんで?縄文色が濃くて良いじゃないかと言っている。
>>292
お前は同和利権ヤクザと混同している。
関西に陰険な同和政治が多いだけで
どちらかといえば関西人に差別された東国人が発端だ。
今の同和とはきっともはや血の繋がりは無い。
なんで?縄文色が濃くて良いじゃないかと言っている。
>>292
お前は同和利権ヤクザと混同している。
関西に陰険な同和政治が多いだけで
どちらかといえば関西人に差別された東国人が発端だ。
今の同和とはきっともはや血の繋がりは無い。
>>295
その説はあり得ない
同和地区は東国にはないんですが・・・無知なの?
東北には部落はないよ
部落の発祥は近畿地方です
近畿地方の部落民が今の部落民です
その場合関西人が差別されるほうで、熊襲差別とは別のものです
関西人は自分達が選民だと勘違いしているようですが(笑) 全く逆です
自分達の顔を見てみれば分かります
関西人は4世紀頃遅れてきた朝鮮系民族であり、侵略者であり、日本人のガンです
まずご自分の顔をご覧になることをおすすめします
その説はあり得ない
同和地区は東国にはないんですが・・・無知なの?
東北には部落はないよ
部落の発祥は近畿地方です
近畿地方の部落民が今の部落民です
その場合関西人が差別されるほうで、熊襲差別とは別のものです
関西人は自分達が選民だと勘違いしているようですが(笑) 全く逆です
自分達の顔を見てみれば分かります
関西人は4世紀頃遅れてきた朝鮮系民族であり、侵略者であり、日本人のガンです
まずご自分の顔をご覧になることをおすすめします
日本
縄文人&弥生人=モンゴロイドとオーストラロイドの混血=アイヌ・熊襲・隼人・沖縄ウチナンチュ
↓
邪馬台国(大和・純粋日本人)
↓
4世紀頃朝鮮から侵略者がくる(偽邪馬台国・九州説はこれ)
↓
邪馬台国を滅ぼし、ヤマト朝廷を樹立し、天皇を詐称する
↓
記紀かきかえ
↓
今の天皇
縄文人&弥生人=モンゴロイドとオーストラロイドの混血=アイヌ・熊襲・隼人・沖縄ウチナンチュ
↓
邪馬台国(大和・純粋日本人)
↓
4世紀頃朝鮮から侵略者がくる(偽邪馬台国・九州説はこれ)
↓
邪馬台国を滅ぼし、ヤマト朝廷を樹立し、天皇を詐称する
↓
記紀かきかえ
↓
今の天皇
大阪人って三度の飯より差別が好きやねん
そやさかい、サントリー前社長が東日本人のことを「熊襲」とか侮蔑してみたり
「文化程度も著しく低い」とかポロッと口すべらせて馬鹿にするわけや
ま〜県民性やからしゃ〜ないわな
未来永劫、大阪人のこの卑しい根性はかわらんわけや
そやさかい、サントリー前社長が東日本人のことを「熊襲」とか侮蔑してみたり
「文化程度も著しく低い」とかポロッと口すべらせて馬鹿にするわけや
ま〜県民性やからしゃ〜ないわな
未来永劫、大阪人のこの卑しい根性はかわらんわけや
ちなみに穢多非人のルーツは天皇家である
在日朝鮮人と部落民はもしかしてルーツが一緒ってことですか?
301 :世界@名無史さん:2012/10/25(木) 07:50:21.95 0
ロシア人スペイン人ブラジル人の教養のなさ
馬鹿さ丸出しの金好き丸出しの女の顔がキモすぎる
第三世界、途上国
まず、ドイツフランスらあたりの女と目つきが明らかに違う
馬鹿さ丸出しの金好き丸出しの女の顔がキモすぎる
第三世界、途上国
まず、ドイツフランスらあたりの女と目つきが明らかに違う
302 :世界@名無史さん:2012/10/25(木) 08:20:37.32 P
{kind=link}
同和地区は条例、規程、告示、訓令、要綱、要領などを見れば解るよ。
例えば新潟県なら、
新潟県同和地区中小企業振興資金融資要綱
http://www.pref.niigata.lg.jp/shogyoshinko/1196093793938.html
てのがあるね。新潟県内に同和地区があるのがわかる。
あとは新潟県の市町村の条例を見れば同和対策の施設を設置する条例があったりするはず。
新発田市隣保館設置条例
http://www1.g-reiki.net/shibata/reiki_honbun/az20004251.html
これらの条例を見ればその同和対策の施設を作る場所が条例で書いてある。
被差別部落は「被差別」+「部落」だから、とある部落に住んでいる人に被差別意識が無いのであれば被差別部落は存在しない。
「自らが置かれた社会的立場の自覚を促す教育」ってのが義務教育にある。それとセットで同和団体や日教組に所属する先生が
同和地区の子供たちに部落差別の当事者である、被差別意識を植え付けるマインドコントロール教育が1970年代からあった。
その結果、同和地区の子供たちが被差別部落出身者の子孫であると自覚するようになり同和地区=被差別部落になってしまった。
例えば新潟県なら、
新潟県同和地区中小企業振興資金融資要綱
http://www.pref.niigata.lg.jp/shogyoshinko/1196093793938.html
てのがあるね。新潟県内に同和地区があるのがわかる。
あとは新潟県の市町村の条例を見れば同和対策の施設を設置する条例があったりするはず。
新発田市隣保館設置条例
http://www1.g-reiki.net/shibata/reiki_honbun/az20004251.html
これらの条例を見ればその同和対策の施設を作る場所が条例で書いてある。
被差別部落は「被差別」+「部落」だから、とある部落に住んでいる人に被差別意識が無いのであれば被差別部落は存在しない。
「自らが置かれた社会的立場の自覚を促す教育」ってのが義務教育にある。それとセットで同和団体や日教組に所属する先生が
同和地区の子供たちに部落差別の当事者である、被差別意識を植え付けるマインドコントロール教育が1970年代からあった。
その結果、同和地区の子供たちが被差別部落出身者の子孫であると自覚するようになり同和地区=被差別部落になってしまった。
「同和地区=被差別部落」ではない
http://plaza.rakuten.co.jp/salsat999/diary/201206060000/
>被差別部落という用語は戦後に作られたもので、それ以前は「特殊部落」とか「細民部落」と呼ばれていました。これらは江戸時代までは
>「穢多村」と呼ばれており、文字通り穢多が居住していた村落のことでしたが、明治4年の解放令によって穢多という身分がなくなり、特殊部落と呼ぶようになったのです。
>一方、同和地区とは、行政が昭和44年(1969年)に同和対策事業の対象として指定した地区のことで、正式な行政用語です。基本的には
>「同和地区=被差別部落」なのですが、被差別部落だったけれども指定されなかった地区もありますし、被差別部落ではないのに指定された地区もあるので、
>必ずしも「同和地区=被差別部落」というわけではありません。ややこしいですね。
こんな感じらしいよ。自然災害で田畑を失ったり住処が沼地になってしまったりして違う土地でコジキのような
生活をするようになった元農民が住んでいた細民部落と呼ばれる部落も同和地区になったケースもあるらしい。
同和関係者の子供たちは同和進学推奨金とかが同和奨学金とか貰って大学にいける。
職業体験とかいうのもあってそれに参加すれば学校給食調理員や市町村の現業職員に推薦してもらえる。
ただ、そのとき部落差別の当事者であると自覚させられる。
だから同和関係者の子供たちは自分を部落民だと言っちゃうし同和地区を被差別部落だと言っちゃう。
http://plaza.rakuten.co.jp/salsat999/diary/201206060000/
>被差別部落という用語は戦後に作られたもので、それ以前は「特殊部落」とか「細民部落」と呼ばれていました。これらは江戸時代までは
>「穢多村」と呼ばれており、文字通り穢多が居住していた村落のことでしたが、明治4年の解放令によって穢多という身分がなくなり、特殊部落と呼ぶようになったのです。
>一方、同和地区とは、行政が昭和44年(1969年)に同和対策事業の対象として指定した地区のことで、正式な行政用語です。基本的には
>「同和地区=被差別部落」なのですが、被差別部落だったけれども指定されなかった地区もありますし、被差別部落ではないのに指定された地区もあるので、
>必ずしも「同和地区=被差別部落」というわけではありません。ややこしいですね。
こんな感じらしいよ。自然災害で田畑を失ったり住処が沼地になってしまったりして違う土地でコジキのような
生活をするようになった元農民が住んでいた細民部落と呼ばれる部落も同和地区になったケースもあるらしい。
同和関係者の子供たちは同和進学推奨金とかが同和奨学金とか貰って大学にいける。
職業体験とかいうのもあってそれに参加すれば学校給食調理員や市町村の現業職員に推薦してもらえる。
ただ、そのとき部落差別の当事者であると自覚させられる。
だから同和関係者の子供たちは自分を部落民だと言っちゃうし同和地区を被差別部落だと言っちゃう。
俺市役所に行くと気持ち悪いオーラや邪念を感じるんだが部落だったのか
感受性強いから吐きそうになる
感受性強いから吐きそうになる
部落は穢多非人ではないってアホか
京都の崇仁地区を見ろ。
京都の崇仁地区を見ろ。
人口激減・高齢少子化――七条部落は、行政的には崇仁地区と呼ばれ、JR京都駅のすぐ東、鴨川のほとりに位置する。
1960年の住宅地区改良法施行後まちづくりが進められてきたが、河川の付け替えに伴う利害対立や人口減少の問題などにより、多くの事業が残されている。
事業の遅れの影響もあり、かつては1万人以上の人口が今は4分の1に激減し、1250世帯2300人。うち65歳以上が700人、70歳以上が520人と高齢化が激しい。
崇仁小学校の生徒は全体でわずか67人(50世帯)と少子化も著しい。
1960年の住宅地区改良法施行後まちづくりが進められてきたが、河川の付け替えに伴う利害対立や人口減少の問題などにより、多くの事業が残されている。
事業の遅れの影響もあり、かつては1万人以上の人口が今は4分の1に激減し、1250世帯2300人。うち65歳以上が700人、70歳以上が520人と高齢化が激しい。
崇仁小学校の生徒は全体でわずか67人(50世帯)と少子化も著しい。
部落は穢多非人ではなかったら、とっくの昔に部落差別はなくなってるはずなんだがねw
穢多非人の他にも夙や茶筅といった雑種賤民もいるよ。
>>296
関西に連れて来られた東国人が差別されて被差別になったのは普通の話。
ありえない、とか断定するのはおかしいのだが。
同和の知識なんてどーでもいーし反論のしつこさがキモいよ。
俺は自分自身が被差別人でも全く構わんが。
被差別人こそ選民です。
頭の狂った関西人は年中差別している変態だから。
差別されたことの方がかっこいいだろ。武士もマムルークも発端は奴隷から。
お前がネチネチと俺に反論すること以外は全て同意だ。
関西に連れて来られた東国人が差別されて被差別になったのは普通の話。
ありえない、とか断定するのはおかしいのだが。
同和の知識なんてどーでもいーし反論のしつこさがキモいよ。
俺は自分自身が被差別人でも全く構わんが。
被差別人こそ選民です。
頭の狂った関西人は年中差別している変態だから。
差別されたことの方がかっこいいだろ。武士もマムルークも発端は奴隷から。
お前がネチネチと俺に反論すること以外は全て同意だ。
美智子妃殿下は被差別人だと言われているね。
ちゃんと歴史を紐解けば、歴史的特質のある者と
大衆から浮く存在とは根底が同じであることがすぐわかる。
戦国武将の正田隼人家は庄屋身分ながら将軍御目見得格。
得川郷と呼ばれる家康の系図の縁地を年貢免除で所領とした郷士で幕末にも活躍がある。
この正田隼人家に繋がると言われるのが日清製粉を興した正田氏。
幕府初期には発祥地の館林に庄田の名で城下被差別地域に居を構える。隣接は浄土真宗と時宗寺。
城下隣接には日清製粉の前身となる醤油工場がある。
正田氏は江戸後期には米問屋として北関東に勢力を持っていた新田系の身分外有力者。
ちゃんと歴史を紐解けば、歴史的特質のある者と
大衆から浮く存在とは根底が同じであることがすぐわかる。
戦国武将の正田隼人家は庄屋身分ながら将軍御目見得格。
得川郷と呼ばれる家康の系図の縁地を年貢免除で所領とした郷士で幕末にも活躍がある。
この正田隼人家に繋がると言われるのが日清製粉を興した正田氏。
幕府初期には発祥地の館林に庄田の名で城下被差別地域に居を構える。隣接は浄土真宗と時宗寺。
城下隣接には日清製粉の前身となる醤油工場がある。
正田氏は江戸後期には米問屋として北関東に勢力を持っていた新田系の身分外有力者。
徳川家康は被差別民出身という説がある。
まあそれは娯楽物だけどね。
可能性としては全然おかしくない。
可能性としては全然おかしくない。
>>310
東国人が関西に連れて来られた、ってシチュエーションがまずありえない。
東国人は関西から追放される一方で、その逆はありえない。
もともと関西にいた縄文弥生人が、南北に分断され追放されたのが、アイヌや琉球民族です。
余談だが、だから東北人と沖縄人は似てるのだ。
東国人が関西に連れて来られた、ってシチュエーションがまずありえない。
東国人は関西から追放される一方で、その逆はありえない。
もともと関西にいた縄文弥生人が、南北に分断され追放されたのが、アイヌや琉球民族です。
余談だが、だから東北人と沖縄人は似てるのだ。
移配された俘囚や佐伯部のことを言ってるんだろ。
東国の他にも薩摩から移配された隼人司もいるよ。
318 :世界@名無史さん:2012/10/26(金) 22:33:47.08 0
トルコのスルタンはヨーロッパの白人処女の奴隷を母とするらしいな。なんかエロイというかエグイよな。敵国の女に皇帝の子供を生ませるという発想が
俘囚や佐伯部は穢多非人ではないだろ
現実の被差別部落の分布とは差異がありすぎる。
現実の被差別部落の分布とは差異がありすぎる。
俘囚や佐伯部は容姿端麗な美人ばかり連れて来たから、エタヒニンとは似ても似つかない。
それって混血させるための政策じゃ?どっちにしろえたひにんとは関係ないよ
322 :世界@名無史さん:2012/10/27(土) 16:33:57.87 0
オスマン家が奴隷を妻とするようになったのは敵に妻を奪われれてもメンツを守るためだったはず
>>315
俘囚のことだよ。
関西人が関西から日本人を追放したのも確かだが、それはその前段階。
野蛮な大陸からの海賊漢寇が日本列島の西部に追いかけたときのこと。
そもそも関西(中華)人が日本に来たとき西国にはほとんど人口が無かった。
東国にしか比重が無かった。
西国人は蝦夷に侵攻してフロンティアを目指してたんだから、
大量の捕虜を畿内に連れて帰っ来てるだろ。
俘囚のことだよ。
関西人が関西から日本人を追放したのも確かだが、それはその前段階。
野蛮な大陸からの海賊漢寇が日本列島の西部に追いかけたときのこと。
そもそも関西(中華)人が日本に来たとき西国にはほとんど人口が無かった。
東国にしか比重が無かった。
西国人は蝦夷に侵攻してフロンティアを目指してたんだから、
大量の捕虜を畿内に連れて帰っ来てるだろ。
なんだか話が通じないヤツがいるな
穢多=賤民
非人=犯罪者
非人=犯罪者
それはえたひにんを現代の同和ヤクザと捉えているか
古来の伝統ある被差別民と捉えているかの違いの問題だ。
俺は現代社会の言葉じゃない以上、
伝統ある被差別民のことと捉えている。
えたひにんは「悪党」のようなものだ。
鎌倉時代の悪党は荘園勢力の地頭層。
古来の伝統ある被差別民と捉えているかの違いの問題だ。
俺は現代社会の言葉じゃない以上、
伝統ある被差別民のことと捉えている。
えたひにんは「悪党」のようなものだ。
鎌倉時代の悪党は荘園勢力の地頭層。
日本史板で自説が受け入れられないからって、世界史板でそれを披露するってかw >被差別民は俘囚説
事実そうなのかも知れないが、根拠が限りなく不足している。
事実そうなのかも知れないが、根拠が限りなく不足している。
日本史板がどうだかとうお前の妄想を含められても困る。
根拠というか捕虜=差別は普通だろ。
日本特有の被差別民なる概念の方が不明瞭なだけで。
根拠というか捕虜=差別は普通だろ。
日本特有の被差別民なる概念の方が不明瞭なだけで。
>>329
日本史板にこの内容のスレがあり、根拠が無いのに延々自説を提示している人がいるんだよw
君は、まあ別の人なのかも知れないけどね。
捕虜が差別されたのはまあ、そうかも知れんが、それが現在まで続いているかは?????
大体、俘囚って言っても、狩猟生活をやめ、日本人的生活を受け入れた蝦夷をも指すだろ?
日本史板にこの内容のスレがあり、根拠が無いのに延々自説を提示している人がいるんだよw
君は、まあ別の人なのかも知れないけどね。
捕虜が差別されたのはまあ、そうかも知れんが、それが現在まで続いているかは?????
大体、俘囚って言っても、狩猟生活をやめ、日本人的生活を受け入れた蝦夷をも指すだろ?
>>330
俺はしらん。誰かが書いた内容についでで書いてるに過ぎん、
俘囚は東北の土豪だな。基本的には移配俘囚は別の話だ。
安倍元総理とかは俘囚出身と言ってたし、
日本の氏族には帰化系とか俘囚系とかのジャンルが存在する。
だから差別はされとらん。
俺はしらん。誰かが書いた内容についでで書いてるに過ぎん、
俘囚は東北の土豪だな。基本的には移配俘囚は別の話だ。
安倍元総理とかは俘囚出身と言ってたし、
日本の氏族には帰化系とか俘囚系とかのジャンルが存在する。
だから差別はされとらん。
俺が思う被差別民というのは
その時代々々に存在する差別される存在について
系譜を紐付けて脈々と受け継がれる文化を持たせようという
外部からの定義によるものじゃないかと理解している。
だから被差別民という括り自体が本当は固定的でなく、
現在のような利権や陰謀が絡むものは
極めて近年の事象を指しているものと理解している。
本来は歴史のひとつに過ぎないただの社会集団。
その時代々々に存在する差別される存在について
系譜を紐付けて脈々と受け継がれる文化を持たせようという
外部からの定義によるものじゃないかと理解している。
だから被差別民という括り自体が本当は固定的でなく、
現在のような利権や陰謀が絡むものは
極めて近年の事象を指しているものと理解している。
本来は歴史のひとつに過ぎないただの社会集団。
被差別民は利権あまーり絡んでないと俺は見ている。
だいたい、2ch草創期から被差別民に関する書き込みが多数あるが、それに対する妨害工作…
コピペで流すとか、無意味なAAを出しまくるとかの行為がほとんど全くない。
これは実際に彼らが困っているせいだと俺は判断している。
被差別民に関してはネット以前だと、宝島社のムックが結構扱っていたよな。参考になった。
だいたい、2ch草創期から被差別民に関する書き込みが多数あるが、それに対する妨害工作…
コピペで流すとか、無意味なAAを出しまくるとかの行為がほとんど全くない。
これは実際に彼らが困っているせいだと俺は判断している。
被差別民に関してはネット以前だと、宝島社のムックが結構扱っていたよな。参考になった。
やれやれ
きちんと書けよ。
暴力団が上前はねていて、そういう人は反論をネットに書かないとかねw
暴力団が上前はねていて、そういう人は反論をネットに書かないとかねw
変な人がいついて困るな
革ジャン着てる人は部落民
革ジャンなら部落が全くない北海道や東北でも売ってるぞ。
馬鹿だな 革ジャン=皮革=部落なんだよ
橋下徹のエピソードを知らないのか?
橋下徹のエピソードを知らないのか?
お前が異様にバカすぎるだけだ。
馬鹿は部落より下だ。
縄文時代人でもそんな判断せんわ。
馬鹿は部落より下だ。
縄文時代人でもそんな判断せんわ。
奈良平安時代ぐらいにはそういう混血政策があったって網野善彦もそう言ってたけど
それが穢多非人の発祥であるとは彼も言っていない
網野善彦ぐらい読んだらどうだ
それが穢多非人の発祥であるとは彼も言っていない
網野善彦ぐらい読んだらどうだ
ということは最初から網野善彦はなにも関係ないんじゃないか。
勝手に引用して勝手に無関係で何やってんだお前。
勝手に引用して勝手に無関係で何やってんだお前。
網野善彦も知らないバカはいい加減にしろ。
うるせーこの部落オタ
それをいうなら部落キチガイだろ
なんでもいいけど穢多は江戸末期の当て字だから。
えたは番太と同じで○太を表すと考えた方が自然だ。無論侮った呼称だ。
非人は非人番や検非することを表す"非"だ。検非違使というのがある。
人に非(あら)ずはウソだ。
考えても見ろ。公的文書に載せる記述だぞ。
そんなふざけた名前があるか。
えたは番太と同じで○太を表すと考えた方が自然だ。無論侮った呼称だ。
非人は非人番や検非することを表す"非"だ。検非違使というのがある。
人に非(あら)ずはウソだ。
考えても見ろ。公的文書に載せる記述だぞ。
そんなふざけた名前があるか。
部落差別の本質も、このメカニズムから一歩も外れることのない単純素朴なものだ。
かつて日本は激しい差別社会であった。おそらく、それは朝鮮百済王国の差別制度がそのまま日本に持ち込まれ、別の発展を遂げたものであろう。
支配階級が機能するためには、対極底辺に被差別階級が必要なのである。
士農工商の身分差別では、農民の憤懣のやり場がないために、さらに、その下に穢多・非人階層を必要とした。
処刑や汚穢処理を強要し、「汚れた人」というイメージを作り出し、権力に対する理不尽の怒りを彼らへの蔑視にすり替えていったのである。
しかし、それでは穢多部落民は、憤懣のやり場をどこに持ってゆけばよいのか? それも、ちゃんと考えられていた。
人でない非人たちには納税義務はなく、このため江戸時代、もっとも裕福な階層こそ穢多だったのだ。
全国穢多総差配であった弾左衛門屋敷は浅草〜吉原の広大な敷地にあり、内部における豪華さは将軍家以上とも謳われたものであり、
そこには京都の貧乏公家から特級美女ばかりが買われてきていた。
また全国の被差別集落の名主たちの裕福さも目を見張るものであり、江戸期を通じて、もっとも貧しさから遠い階層であった。
さらに、牛馬解体を一手に引き受け、鎌倉仏教による肉食禁止のタンパク質不足のなかで、唯一恵まれた供給源があった。
かつて日本は激しい差別社会であった。おそらく、それは朝鮮百済王国の差別制度がそのまま日本に持ち込まれ、別の発展を遂げたものであろう。
支配階級が機能するためには、対極底辺に被差別階級が必要なのである。
士農工商の身分差別では、農民の憤懣のやり場がないために、さらに、その下に穢多・非人階層を必要とした。
処刑や汚穢処理を強要し、「汚れた人」というイメージを作り出し、権力に対する理不尽の怒りを彼らへの蔑視にすり替えていったのである。
しかし、それでは穢多部落民は、憤懣のやり場をどこに持ってゆけばよいのか? それも、ちゃんと考えられていた。
人でない非人たちには納税義務はなく、このため江戸時代、もっとも裕福な階層こそ穢多だったのだ。
全国穢多総差配であった弾左衛門屋敷は浅草〜吉原の広大な敷地にあり、内部における豪華さは将軍家以上とも謳われたものであり、
そこには京都の貧乏公家から特級美女ばかりが買われてきていた。
また全国の被差別集落の名主たちの裕福さも目を見張るものであり、江戸期を通じて、もっとも貧しさから遠い階層であった。
さらに、牛馬解体を一手に引き受け、鎌倉仏教による肉食禁止のタンパク質不足のなかで、唯一恵まれた供給源があった。
なんだこいつ解放同盟かw典型的な解同論理じゃん
解同のいう江戸起源説は明らかな間違いで否定されているのにね
解同のいう江戸起源説は明らかな間違いで否定されているのにね
解同氏ねよw
まあ必ずしも間違いではないしコピペだと思うけどステレオタイプがいくつも混ざってるよ。
士農工商は今ではほぼ実体無しと言われているし当然その下のえたひにんも。
上か下かではなく社会には生産層と支配層、その他がある。
今で言うサービス業、あと実務公務員、これらが身分外。
えたの最も重要な仕事は警察だよ。
平和な日本で生活が苦しい時は当然これらの職業は民衆に嫌われる。
私服警官なんてうんざりだから見た目でわかるようにして欲しいと訴える。
当時は全国統一の警察なんて無かったから各藩でそれぞれ雇われていた百姓以外の者。
しかも警察とか公務員って親の職業を大体受け継ぐんだよな。
士農工商は今ではほぼ実体無しと言われているし当然その下のえたひにんも。
上か下かではなく社会には生産層と支配層、その他がある。
今で言うサービス業、あと実務公務員、これらが身分外。
えたの最も重要な仕事は警察だよ。
平和な日本で生活が苦しい時は当然これらの職業は民衆に嫌われる。
私服警官なんてうんざりだから見た目でわかるようにして欲しいと訴える。
当時は全国統一の警察なんて無かったから各藩でそれぞれ雇われていた百姓以外の者。
しかも警察とか公務員って親の職業を大体受け継ぐんだよな。
古代末から中世にかけて、一般農民の共同体からはじき出され、あるいは脱落・流出し、
中世非人(社会外の社会)となった人びとが、宿・声聞師・河原者などの賤民集団を形成し、
河原者はケガレの処理を中心とする職能集団として、さまざまな仕事を担う。
そして、動物の死骸などケガレの処理は、本来、河原などに埋めてしまえばこと足りるのだが、
病弱になって働けなくなった耕作用の牛馬は、単に飼い置くわけにもいかず、その屠殺も河原者の仕事である。
こうしてできた動物の死骸から様々な物を作る化製業系の仕事が発生する。
皮革の製造はいうに及ばず、接着剤としての膠や漢方薬である牛黄など、さまざまなものを工夫して製造した。
戦国大名が、城下町築造の際に、自分の出身地から皮多を引き連れて来て優遇したのは、こうした特殊技能を重視してのことである。
また、犯罪の取り締まりや処罰も、ケガレの処理の一形態で、古くから河原者もたずさわっていた。
近代になっても、各地の部落に十手や御用提灯が保存されていたのは、そうしたことに由来する。
また、ケガレの処理から発展したものに、権門の住居のトータルコーディネーターという役割もある。
本来は、庭を清浄に保つのがケガレ処理の本義であろうが、徐々に土木技術や作庭技術・理論を発展させていった。
また、警察業務との関連もあるが、庭者の陰の任務として、諜報活動も重要な仕事であった。
こうしたケガレ処理関連の仕事とは別に、河原者たちは営々と農地の開墾・耕作に努力し、江戸時代に入ってからは、
一般の百姓とかなり近い存在となっている。
中世非人(社会外の社会)となった人びとが、宿・声聞師・河原者などの賤民集団を形成し、
河原者はケガレの処理を中心とする職能集団として、さまざまな仕事を担う。
そして、動物の死骸などケガレの処理は、本来、河原などに埋めてしまえばこと足りるのだが、
病弱になって働けなくなった耕作用の牛馬は、単に飼い置くわけにもいかず、その屠殺も河原者の仕事である。
こうしてできた動物の死骸から様々な物を作る化製業系の仕事が発生する。
皮革の製造はいうに及ばず、接着剤としての膠や漢方薬である牛黄など、さまざまなものを工夫して製造した。
戦国大名が、城下町築造の際に、自分の出身地から皮多を引き連れて来て優遇したのは、こうした特殊技能を重視してのことである。
また、犯罪の取り締まりや処罰も、ケガレの処理の一形態で、古くから河原者もたずさわっていた。
近代になっても、各地の部落に十手や御用提灯が保存されていたのは、そうしたことに由来する。
また、ケガレの処理から発展したものに、権門の住居のトータルコーディネーターという役割もある。
本来は、庭を清浄に保つのがケガレ処理の本義であろうが、徐々に土木技術や作庭技術・理論を発展させていった。
また、警察業務との関連もあるが、庭者の陰の任務として、諜報活動も重要な仕事であった。
こうしたケガレ処理関連の仕事とは別に、河原者たちは営々と農地の開墾・耕作に努力し、江戸時代に入ってからは、
一般の百姓とかなり近い存在となっている。
江戸時代の中期以降は、町人文化の発展に連動して、製造業、とりわけ雪踏の裏張りの原料である皮革を独占的に製造していた関係で、
履物関連産業が一挙に開花・発展して、皮多村の経済が飛躍的発展をとげる。
また、動物の屠殺や化製業に関する知識、および処刑などの刑吏役から来る人体に関する知識が発展して、
医療技術にたけた穢多身分の人が輩出される。
1777年(安永6)、武州榛沢郡新戒村の穢多が「医道巧者」なので、もっと活躍してもらおうとの世論が高まり、
村や近郷の人々あげての身分引き上げ嘆願運動があったことは、あまりに有名である。
また、1771年(明和8)の小塚原で杉田玄白らに人体解剖をしてみせたのが、刑場で働く穢多の虎松のお祖父さんであった。
また、京都府蓮台野村の益井元右衛門は眼病院を設立して、相当高度の外科手術をし、
果ては部落の子どもたちにドイツ語まで教えていた。
他にも多くの地域で被差別部落から名医が多く出ているが、これらは斃牛馬処理や刑吏役と強い結びつきを想起させる。
履物関連産業が一挙に開花・発展して、皮多村の経済が飛躍的発展をとげる。
また、動物の屠殺や化製業に関する知識、および処刑などの刑吏役から来る人体に関する知識が発展して、
医療技術にたけた穢多身分の人が輩出される。
1777年(安永6)、武州榛沢郡新戒村の穢多が「医道巧者」なので、もっと活躍してもらおうとの世論が高まり、
村や近郷の人々あげての身分引き上げ嘆願運動があったことは、あまりに有名である。
また、1771年(明和8)の小塚原で杉田玄白らに人体解剖をしてみせたのが、刑場で働く穢多の虎松のお祖父さんであった。
また、京都府蓮台野村の益井元右衛門は眼病院を設立して、相当高度の外科手術をし、
果ては部落の子どもたちにドイツ語まで教えていた。
他にも多くの地域で被差別部落から名医が多く出ているが、これらは斃牛馬処理や刑吏役と強い結びつきを想起させる。
江戸時代の身分制度が発祥なら、江戸幕府などの何らかの文書が残っているはずだが、何も
残っていません
残っていません
穢多非人=コサックみたいなものだろ
部落論はもはや知識の洪水で特殊な学問体系と化しているからね。
ケガレの仕事というがそれって清掃業務じゃないの。
言うことがいちいち大袈裟だよな。
きっかけは廃藩置県で全国の公務職が全員失職したことだから。
貧困は差別を生む。またこの時の没落士族も同じ状態。
坊ちゃんに出て来る乳母も士族だし遊郭に売られた娘もたくさんいた。
ただ文化の発信源の近畿ではそういう逆転身分は少なかったから
相対性が失われたのだろう。
ケガレの仕事というがそれって清掃業務じゃないの。
言うことがいちいち大袈裟だよな。
きっかけは廃藩置県で全国の公務職が全員失職したことだから。
貧困は差別を生む。またこの時の没落士族も同じ状態。
坊ちゃんに出て来る乳母も士族だし遊郭に売られた娘もたくさんいた。
ただ文化の発信源の近畿ではそういう逆転身分は少なかったから
相対性が失われたのだろう。
コサックの語源はトルコ語の自由人という意味で
本来はツアーリの尖兵ではなかった。
プガチョフなど公然とツアーリに反逆し、
さらに他民族で異教徒のチュバシ族などと連携するなど
どちらかというとロシア帝国内の不穏分子だった。
本来はツアーリの尖兵ではなかった。
プガチョフなど公然とツアーリに反逆し、
さらに他民族で異教徒のチュバシ族などと連携するなど
どちらかというとロシア帝国内の不穏分子だった。
部落の貧困化は差別問題とはまったく別のところからやってきたことにある。
それが、松方デフレ政策にほかならない。
1877年(明治10)に勃発した西南戦争で、明治政府は最強のプロ戦闘集団である薩摩武士を相手に多額の軍費を使い、
不換紙幣を乱発したために、悪性のインフレに見舞われた。
その解決のために、松方正義大蔵卿が急激な紙幣整理というハードランディング方式をとったために、
一挙にデフレになり、部落の製造業が壊滅的打撃を受けたのである。
決して、部落が狙い撃ちされて被害をこうむったわけではなく、また差別されて貧乏になったわけでもない。
それが、松方デフレ政策にほかならない。
1877年(明治10)に勃発した西南戦争で、明治政府は最強のプロ戦闘集団である薩摩武士を相手に多額の軍費を使い、
不換紙幣を乱発したために、悪性のインフレに見舞われた。
その解決のために、松方正義大蔵卿が急激な紙幣整理というハードランディング方式をとったために、
一挙にデフレになり、部落の製造業が壊滅的打撃を受けたのである。
決して、部落が狙い撃ちされて被害をこうむったわけではなく、また差別されて貧乏になったわけでもない。
コサックは遊牧トルコ系の軍事技術が
ウクライナ等の没落貴族を取り込んだ形で生まれた傭兵身分だね。
キリスト教改宗トルコ人から姿形を変えて原型も消したもの。
明治警察を従来通り分権行政とするか集権行政とするかは、
意見の分かれたところだったが、結局集権警察一本となった。
これで現地の行政官えたは士族とともに失職。
代わりに来たのが大久保派の川路利良率いる警察集団。
薩摩旧士族を中心とした彼らの蔑称は薩摩っぽ。今でいうマッポ。
ウクライナ等の没落貴族を取り込んだ形で生まれた傭兵身分だね。
キリスト教改宗トルコ人から姿形を変えて原型も消したもの。
明治警察を従来通り分権行政とするか集権行政とするかは、
意見の分かれたところだったが、結局集権警察一本となった。
これで現地の行政官えたは士族とともに失職。
代わりに来たのが大久保派の川路利良率いる警察集団。
薩摩旧士族を中心とした彼らの蔑称は薩摩っぽ。今でいうマッポ。
コサックは逃亡小作人が自治を行うようになったもの。
世界史板なのに何故か日本史の話が多いな?
穢多非人が本来差別されてないなら、なんでいまだに差別がなくならないんだ?
答えろバカ
主張が矛盾しているのに気付かないのか?
答えろバカ
主張が矛盾しているのに気付かないのか?
現在の部落差別の起源は、関ヶ原や大阪の陣の後、徳川方が豊臣方を差別して、 土地や財産を奪って
被差別部落に押し込めたのが起源
被差別部落に押し込めたのが起源
部落民が裕福なんてあり得ません
街道うざ
うざかろうが、何だろうが2ch過去ログ見ていると、部落に対する差別書き込みに対して
組織的な対抗書き込みがほぼ皆無なことを考えると、経済状況が自ずと分かる。
組織的な対抗書き込みがほぼ皆無なことを考えると、経済状況が自ずと分かる。
牛や馬なんか昔の人は食べないだろ。
ヨーロッパ人がなんで容姿淡麗か
閑話休題
いまだに差別がなくならないというより今から始まった差別なんだな。明治頃に。
コサックは一般にスラブ人流浪者と言われるね。
しかしやはり原型はトルコと言わざるを得ないのでは。
コサックは一般にスラブ人流浪者と言われるね。
しかしやはり原型はトルコと言わざるを得ないのでは。
江戸時代って言ってたじゃんw
悪銭身につかず、って言うだろ
簡単にできたものは簡単になくなるはずだが
簡単にできたものは簡単になくなるはずだが
実際障碍者差別も、女性差別もなくなってるのに部落差別だけなくならないのはなんで?
江戸?明治?どっちにしろ歴史が浅いならすぐになくなるはずだけど?
市松人形よりも昔のフランス人形のほうが可愛いよね
ビスクドールで検索してみ、当時の細目の市松よりも
はるかに可愛い人形がたくさん出てくるから
白人の子供がモチーフなだけに凄く可愛い
ビスクドールで検索してみ、当時の細目の市松よりも
はるかに可愛い人形がたくさん出てくるから
白人の子供がモチーフなだけに凄く可愛い
だから江戸時代はウソだって。後期の頃ならあったかも。
少なくとも政治が設定したとかはウソ。
フランス人形(こわいんだけど)
http://www.iza.ne.jp/images/user/20090623/527878.jpg
ひな人形
http://img5.blogs.yahoo.co.jp/ybi/1/dc/f3/kyonko_happy_life/folder/433235/img_433235_3291567_0
少なくとも政治が設定したとかはウソ。
フランス人形(こわいんだけど)
http://www.iza.ne.jp/images/user/20090623/527878.jpg
{kind=link}
ひな人形
http://img5.blogs.yahoo.co.jp/ybi/1/dc/f3/kyonko_happy_life/folder/433235/img_433235_3291567_0
無くならないってことは、穢多非人は少なくとも平安時代からあって、賤民だってことだ。
うんうん。
>>379
つまり穢多非人は朝鮮人ってことですね
つまり穢多非人は朝鮮人ってことですね
無くなるかというより今が最大のブームだから。
平安時代にそんなものは無い。こじつけ
平安時代にそんなものは無い。こじつけ
>>380
朝廷での権力闘争に敗れた百済系末裔が多いのでは?
朝廷での権力闘争に敗れた百済系末裔が多いのでは?
西原理恵子を見れば分かるだろ
橋下徹でいいじゃん
385 :世界@名無史さん:2012/11/03(土) 08:57:17.50 0
野中広務
松本龍
山城新伍
橋下徹
西原理恵子
松本龍
山城新伍
橋下徹
西原理恵子
>>380,382
冗談はさておき明治時代の定説だな。
被差別階層に帰化人がいた可能性は十分に高いが、
えた非人という役職は当時無い。
時の身分離反者が時代ごとに積み重ねて作られた階層だから、
流動し過ぎて系統は言い切れない。
冗談はさておき明治時代の定説だな。
被差別階層に帰化人がいた可能性は十分に高いが、
えた非人という役職は当時無い。
時の身分離反者が時代ごとに積み重ねて作られた階層だから、
流動し過ぎて系統は言い切れない。
今の在日と現象的に一緒でしょ。
穢多非人=古代の在日朝鮮人と捉えたほうが、すべてうまく説明できる。
だからと言って正しいとは限らないが。
穢多非人=古代の在日朝鮮人と捉えたほうが、すべてうまく説明できる。
だからと言って正しいとは限らないが。
違うでしょ。
古代の在日はマジョリティで権力者だから。
もちろん関西での話だが、
その中にも貴賎両者が存在したので
弱者は異郷では差別されただろうという話。
古代の在日はマジョリティで権力者だから。
もちろん関西での話だが、
その中にも貴賎両者が存在したので
弱者は異郷では差別されただろうという話。
違わないじゃんw
現在の在日がわからんからわからんわ。
じゃあもう違わないでよい。
じゃあもう違わないでよい。
こいつらの話聞いてても全然勉強にならん
真の学問とは、聞いて「あ!そうか」とすっきりしてモヤモヤが晴れる感じなんだが
真の学問とは、聞いて「あ!そうか」とすっきりしてモヤモヤが晴れる感じなんだが
理屈こねまわしてるだけだもんな〜
関東→近年狭山事件とかで問題になるまでは、あまり気にされる事もなく共存していた。穢多頭の弾左衛門や、
非人頭の車善七といった、江戸幕府から特権を得られていた部落民も存在していた
関西→徹底した蔑視の対象とされていた。
非人頭の車善七といった、江戸幕府から特権を得られていた部落民も存在していた
関西→徹底した蔑視の対象とされていた。
関西人は差別が好っきゃねん
新撰組は穢多
やっぱ好っきゃね〜ん
火消しの「め組」や岡っ引きも穢多
車善七というと寅さんのモデルだね。
弾左衛門は鎌倉の由比ガ浜の長吏。
もともと関東は小田原の太郎左衛門という長吏頭が有力だったのだが、
家康入府で関東支配層が入れ替わり弾左衛門が関東長吏頭を引き継いだ。
江戸にはその他品川の松右衛門など有力者が揃う。
元々町奴と旗本奴の抗争など下町気風の強い江戸。
これらが後年火消しや岡っ引や歌舞伎役者の土台となっていく。
町奴幡随院長兵衛は元武士の子。旗本奴水野十郎左衛門は名門の出。
傾奇者の系譜が脈々と継がれるのは東国のお国柄。
官軍支援した清水次郎長や国定忠治、自由民権運動秩父事件のリーダー・侠客の田代栄助。
新撰組、そして母体八王子千人同心は子孫がいまも郷士身分を主張している。
かれらの仕事は捕吏捕縛等の半武士半農。いわゆる長吏。関西風にいえばえた。
弾左衛門は鎌倉の由比ガ浜の長吏。
もともと関東は小田原の太郎左衛門という長吏頭が有力だったのだが、
家康入府で関東支配層が入れ替わり弾左衛門が関東長吏頭を引き継いだ。
江戸にはその他品川の松右衛門など有力者が揃う。
元々町奴と旗本奴の抗争など下町気風の強い江戸。
これらが後年火消しや岡っ引や歌舞伎役者の土台となっていく。
町奴幡随院長兵衛は元武士の子。旗本奴水野十郎左衛門は名門の出。
傾奇者の系譜が脈々と継がれるのは東国のお国柄。
官軍支援した清水次郎長や国定忠治、自由民権運動秩父事件のリーダー・侠客の田代栄助。
新撰組、そして母体八王子千人同心は子孫がいまも郷士身分を主張している。
かれらの仕事は捕吏捕縛等の半武士半農。いわゆる長吏。関西風にいえばえた。
こうゆうのはカムイ伝なんかを読むとよい。
第一部は勉強にはなるがひたすら絶望の嵐。多大にプロパガンダを含むが為にはなる。
第二部は舞台を江戸に移し一転してカラッとした冒険活劇になる。
第一部を関西、第二部を関東とよむとわかりやすい。
第一部は勉強にはなるがひたすら絶望の嵐。多大にプロパガンダを含むが為にはなる。
第二部は舞台を江戸に移し一転してカラッとした冒険活劇になる。
第一部を関西、第二部を関東とよむとわかりやすい。
400
人生難易度
ultra easy 美人
very easy イケメン
easy 普通女
normal フツメン
hard ブサメン
very hard キモメン
ultra hard ブス
ultra easy 美人
very easy イケメン
easy 普通女
normal フツメン
hard ブサメン
very hard キモメン
ultra hard ブス
容姿格差
http://livedoor.blogimg.jp/hyukkyyy_/imgs/4/1/4162af01.jpg
http://sonnnamono.up.seesaa.net/image/8d357030.jpg
http://www.sanspo.com/baseball/images/20121001/car12100105030000-p2.jpg
http://livedoor.blogimg.jp/hyukkyyy_/imgs/4/1/4162af01.jpg
{kind=link}
http://sonnnamono.up.seesaa.net/image/8d357030.jpg
{kind=link}
http://www.sanspo.com/baseball/images/20121001/car12100105030000-p2.jpg
{kind=link}
一重がいい・一重が進化してるってのは全部ステマだな
平安美人は全部一重ってのもステマ
平安美人は全部一重ってのもステマ
一重まぶたは二重まぶたが進化したものなのですか?
http://detail.chiebukuro.yahoo.co.jp/qa/question_detail/q1335302668
http://detail.chiebukuro.yahoo.co.jp/qa/question_detail/q1335302668
寒さに適応しただけであって進化ではない
順番にいうと一重が進化は一応正しい。
猿はまぶたのヒダが無い二重だから。後発であることは確実。
進化=上位とは限らないだけ。
こんな変な顔のヤギだって進化によるものだ。
http://livedoor.blogimg.jp/laba_q/imgs/c/a/cad86d65.jpg
猿はまぶたのヒダが無い二重だから。後発であることは確実。
進化=上位とは限らないだけ。
こんな変な顔のヤギだって進化によるものだ。
http://livedoor.blogimg.jp/laba_q/imgs/c/a/cad86d65.jpg
{kind=link}
"一重がいいはおかしい" ってのもどうかな。
一般に写真や映像で人物判断するようになったのは極めて近年のことだから。
人間には"たたずまい"というものがあって、
落ち着き方や声なども姿形に含まれる。
一重でも二重でも印象の良い人物はこれらが備わる。
生活のほとんどは顔なんて見ない。
二重は、顔の良し悪しというより、
目に力のある人ほど先の好印象の性質を持ちやすいから。
平安美人は一重はステマ
一般に写真や映像で人物判断するようになったのは極めて近年のことだから。
人間には"たたずまい"というものがあって、
落ち着き方や声なども姿形に含まれる。
一重でも二重でも印象の良い人物はこれらが備わる。
生活のほとんどは顔なんて見ない。
二重は、顔の良し悪しというより、
目に力のある人ほど先の好印象の性質を持ちやすいから。
平安美人は一重はステマ
408 :世界@名無史さん:2012/11/17(土) 14:07:01.85 0
有色人趣が優雅な生活をし、科学軍事的にも
白人趣を圧倒する立場をしていたなら、
有色人種の容姿を美しいと思うだろう。
モンゴルがヨーロッパに進出したときには
圧倒的に強いモンゴル人の容姿の特徴を真似する者が
ヨーロッパ人たちに流行したという記録がある。
美に引かれるというものはその背後にある、自己が受容したいと
いう存在に憧れるのだ。
白人趣を圧倒する立場をしていたなら、
有色人種の容姿を美しいと思うだろう。
モンゴルがヨーロッパに進出したときには
圧倒的に強いモンゴル人の容姿の特徴を真似する者が
ヨーロッパ人たちに流行したという記録がある。
美に引かれるというものはその背後にある、自己が受容したいと
いう存在に憧れるのだ。
そう。結局、背景に経済的、軍事的優位性がある。世界をそうやって制覇した人たち=美しい
ということ。
壮大なステマ?その通り。だが歴史はそれが繰り返されてきた。
ということ。
壮大なステマ?その通り。だが歴史はそれが繰り返されてきた。
410 :世界@名無史さん:2012/11/17(土) 15:23:49.28 0
餓死寸前の女性がアイドルやったり、グラビアやAVに出たりしたのは見た事ないけど、仮にそんなのがあっても
絶対抜けないはずです。ガリガリ過ぎるとボインがなくなり、性欲を起こす女らしいラインも消えるからです。
美人が美人でいられるのは単に生まれつきの容姿だけではなくて、文化や経済のお陰でもあるんです。
日本が経済大国になったからこそ世界一の美人も鏡よ鏡世界一番カワイイになった訳です。
絶対抜けないはずです。ガリガリ過ぎるとボインがなくなり、性欲を起こす女らしいラインも消えるからです。
美人が美人でいられるのは単に生まれつきの容姿だけではなくて、文化や経済のお陰でもあるんです。
日本が経済大国になったからこそ世界一の美人も鏡よ鏡世界一番カワイイになった訳です。
壮大なステマだけじゃねえけどな。
白人の自己管理には見習うべきものがある。
他人の印象を当然考慮しているよ。
黄色人種にとっては優先度が低いだけで。
目が大きければ動物界普遍的に影響するし。
ただ、それが全てだといいたがる謎の白人原理主義が意味不明なだけであって。
白人の自己管理には見習うべきものがある。
他人の印象を当然考慮しているよ。
黄色人種にとっては優先度が低いだけで。
目が大きければ動物界普遍的に影響するし。
ただ、それが全てだといいたがる謎の白人原理主義が意味不明なだけであって。
>目が大きければ動物界普遍的に影響するし。
これには根拠がないな。
これには根拠がないな。
413 :世界@名無史さん:2012/11/17(土) 16:24:45.56 0
>>411 そこは「白人」というより「女性」でしょ。マスコミには明らか変な整形原理主義いるよな。
韓国がまた日本や欧米以上に多い。日本の埼玉美人上位8神が韓国人や白人に負ける気はしないが。
日本人は埼玉だけでも綺麗な白人に勝つ自信は100%あるけどな。日本美人世界無双
韓国がまた日本や欧米以上に多い。日本の埼玉美人上位8神が韓国人や白人に負ける気はしないが。
日本人は埼玉だけでも綺麗な白人に勝つ自信は100%あるけどな。日本美人世界無双
目玉の動物避けとかあるだろ。
農家に行け。
農家に行け。
デカい目玉みたいな模様で外敵を威嚇する昆虫なら沢山いるな
埼玉なんぞ島根にも負ける。
整形にも悪くない部分もある。
外形を変えてツボが刺激されるとかがあるなら、内面にも効果もあるだろう。
アスリートの肉体改造なんてもっとなんでもやる。
理由が外見目的ではないだけであって。
ただ外見至上主義者なんて、果ては2次元アニオタレベルまで存在するから、
キリがなくて意味が無い。
整形にも悪くない部分もある。
外形を変えてツボが刺激されるとかがあるなら、内面にも効果もあるだろう。
アスリートの肉体改造なんてもっとなんでもやる。
理由が外見目的ではないだけであって。
ただ外見至上主義者なんて、果ては2次元アニオタレベルまで存在するから、
キリがなくて意味が無い。
>>408
その記録を教えてくれ
圧倒的に強いも何もポーランドには防がれたし、実質ロシアぐらいしか侵せていないはずだが
トルコ人がバルカンを震撼させたときはみな赤褐色の肌に憧れたのか?
ケルト人・ゲルマン人は黒髪短矩のローマ人に憧れたのか?
少なくともフン族がローマを劫掠した時は醜いと受け止められていたな
その記録を教えてくれ
圧倒的に強いも何もポーランドには防がれたし、実質ロシアぐらいしか侵せていないはずだが
トルコ人がバルカンを震撼させたときはみな赤褐色の肌に憧れたのか?
ケルト人・ゲルマン人は黒髪短矩のローマ人に憧れたのか?
少なくともフン族がローマを劫掠した時は醜いと受け止められていたな
>>417
以前Eテレで見たな。草原を馬で移動するには、短躯の方が有利。長距離を移動できるしな。
以前Eテレで見たな。草原を馬で移動するには、短躯の方が有利。長距離を移動できるしな。
貧しい国だとデブの方がモテるんだってな
90年代の東南アジアの俳優にほっぺたがプックリしたコデブが多いし
90年代の東南アジアの俳優にほっぺたがプックリしたコデブが多いし
今年の夏、日本一厳しい登山道と言われている南八甲田の山に登ってきた。
元は自動車道として作られたので、傾斜はたいしたコトがないのだが、とにかく凄い藪と、地面がやたら荒れて
いることに閉口した。背を低くして、延々藪の中の道を踏破するつらさ。
ジャングルで米軍が敗れた理由がなんとなく分かった。
また、こういったトコを踏破して、タケノコを採るみたいなのを競技化すると、日本人に無茶有利だとも思ったw
現在のオリンピック競技って、大抵背が高い方が有利な競技ばかり…。不公平だなw
元は自動車道として作られたので、傾斜はたいしたコトがないのだが、とにかく凄い藪と、地面がやたら荒れて
いることに閉口した。背を低くして、延々藪の中の道を踏破するつらさ。
ジャングルで米軍が敗れた理由がなんとなく分かった。
また、こういったトコを踏破して、タケノコを採るみたいなのを競技化すると、日本人に無茶有利だとも思ったw
現在のオリンピック競技って、大抵背が高い方が有利な競技ばかり…。不公平だなw
外見から受ける感覚は条件反射による。
商品のロゴを見せられるとともに便利であった経験が伴うと、
そのロゴを見るだけで快適のイメージと結び付く。
これが代々積み重なれば染色体情報に組み込まれ本能となり、
数代だけだと文化による後天的経験則。
だから白人の外見優越というのは無根拠では有り得ないが、
サーベルタイガーの例もあるように正しいとも限らない。
自然淘汰の世界では、機能や天候などへの環境順応性の方が遥かに重要。
商品のロゴを見せられるとともに便利であった経験が伴うと、
そのロゴを見るだけで快適のイメージと結び付く。
これが代々積み重なれば染色体情報に組み込まれ本能となり、
数代だけだと文化による後天的経験則。
だから白人の外見優越というのは無根拠では有り得ないが、
サーベルタイガーの例もあるように正しいとも限らない。
自然淘汰の世界では、機能や天候などへの環境順応性の方が遥かに重要。
その通りだが、今後その環境がどう変わるか分からない以上、何が優れているかも変動し将来の
優れているモノの予測もできない。
ま、生物多様性はだから大切だとも言える。
優れているモノの予測もできない。
ま、生物多様性はだから大切だとも言える。
ここでの議論は、染色体情報より後天的近代プロパガンダの影響の方が強いだろう、
という話だと認識している。
白人優越性を主張する者は、実物と接するでもなく、
思考の交換もなく、ただメディアによる作られた動画・静止画により、
結論付けているだけなので、本能に基づいたものでは有り得ない。
という話だと認識している。
白人優越性を主張する者は、実物と接するでもなく、
思考の交換もなく、ただメディアによる作られた動画・静止画により、
結論付けているだけなので、本能に基づいたものでは有り得ない。
ハインリヒヒムラー
「この戦いは国家社会主義の戦いであり、我がゲルマンの、
北方人種の高貴な血に基づいた世界観のための戦いである。
だが我々と別に 〜略〜 発音しにくい名前を持つ体格の悪い連中だ。
そんな連中は憐れみをかけずに撃ち殺せばよい。
こうした人間どもは、ユダヤという一つの宗教と共産主義という
一つのイデオロギーの下に統合されている。
諸君らが東方で戦っている相手はこうした人間以下の劣等人種なのである。
ある時はフン族、またある時はマジャール、さらにはタタール、
チンギス・ハン、モンゴルと。名前を変えても劣等人種に変わりはない。」
この時代は「演説」という手法が音声メディアで広がって、
その技術が民衆を心酔させた。
結局は新技術に魅了されたものなので価値観判断を見誤った。
「この戦いは国家社会主義の戦いであり、我がゲルマンの、
北方人種の高貴な血に基づいた世界観のための戦いである。
だが我々と別に 〜略〜 発音しにくい名前を持つ体格の悪い連中だ。
そんな連中は憐れみをかけずに撃ち殺せばよい。
こうした人間どもは、ユダヤという一つの宗教と共産主義という
一つのイデオロギーの下に統合されている。
諸君らが東方で戦っている相手はこうした人間以下の劣等人種なのである。
ある時はフン族、またある時はマジャール、さらにはタタール、
チンギス・ハン、モンゴルと。名前を変えても劣等人種に変わりはない。」
この時代は「演説」という手法が音声メディアで広がって、
その技術が民衆を心酔させた。
結局は新技術に魅了されたものなので価値観判断を見誤った。
425 :世界@名無史さん:2012/11/17(土) 22:04:38.24 0
白人の欠点:毛深い ガタイがデカイ デブりやすい 腋臭らしい 白すぎて血管が青く見える人がいる
東アジア系の欠点:顔がのっぺりしすぎ 目が一重
東南アジア系の欠点:色が黒すぎて貧乏臭い
欧州美人+東アジア美人+東南アジア美人=日本美人=3人種の容姿の良さを生かし、
欠点は足して3で割った、肌の白さ黒さ具合、目鼻立ちと唇、胸の大きさ、爪先までの奇跡的黄金比=完璧
東アジア系の欠点:顔がのっぺりしすぎ 目が一重
東南アジア系の欠点:色が黒すぎて貧乏臭い
欧州美人+東アジア美人+東南アジア美人=日本美人=3人種の容姿の良さを生かし、
欠点は足して3で割った、肌の白さ黒さ具合、目鼻立ちと唇、胸の大きさ、爪先までの奇跡的黄金比=完璧
426 :世界@名無史さん:2012/11/17(土) 23:43:53.94 0
言うだけ言って後から矛盾だらけになるのは、
いつの時代のネトウヨも同じなのだ。
だから後付けのつじつま合わせが大変だ。
いつの時代のネトウヨも同じなのだ。
だから後付けのつじつま合わせが大変だ。
428 :世界@名無史さん:2012/11/18(日) 00:38:53.18 0
ベラルーシ人は美しいな。下のなんて一般人なのにアメリカでだったらトップクラスの美形に入るんじゃないか。ハリウッド映画とかに出てくる女優レベル
http://boss.allfine.jp/wp-content/uploads/2012/05/%E3%83%99%E3%83%A9%E3%83%AB%E3%83%BC%E3%82%B7.jpg
http://peaceman.img.jugem.jp/20080509_316004.jpg
http://livedoor.blogimg.jp/akiravich/imgs/e/3/e30a7767.JPG
http://livedoor.blogimg.jp/akiravich/imgs/9/9/993378e9.JPG
http://boss.allfine.jp/wp-content/uploads/2012/05/%E3%83%99%E3%83%A9%E3%83%AB%E3%83%BC%E3%82%B7.jpg
{kind=link}
http://peaceman.img.jugem.jp/20080509_316004.jpg
{kind=link}
http://livedoor.blogimg.jp/akiravich/imgs/e/3/e30a7767.JPG
{kind=link}
http://livedoor.blogimg.jp/akiravich/imgs/9/9/993378e9.JPG
{kind=link}
正直フンの血は全員入っとるわ
ところが、東洋人の形質って、西洋人の血に混じると急速になくなっていくんだよな。
なんとも。
なんとも。
東洋人が力不足なので形質を持つ者は
小さくなって暮らしてるだけだ。
日帝が世界支配とかをしていたら、
今頃東欧では東洋人顔だらけだ。
小さくなって暮らしてるだけだ。
日帝が世界支配とかをしていたら、
今頃東欧では東洋人顔だらけだ。
433 :世界@名無史さん:2012/11/19(月) 12:08:24.67 0
>>432 トルコ系民族は力を持っていたが、白人化した。タタール人とロシア人の区別はつかない。
ハザール人やタタール人は長い間征服者だったのだ。カルムイクのように東洋人顔を残したければ
イラン系やスラブ系との結婚は避けねばならない。形質は黒人が一番強く白人がその次で、黄人は一番弱い。
でも立場的には黒人が一番弱かった。最近は芸能・スポーツのトップや閣僚になる人も増えてきたが、せいぜい70年代以後の話。
実際は混血は白黄でも黒黄でも、純粋な人種よりモンゴロイド(ーOー)的な薄さが加わりマイルドな風味になるがな。東亜系=顔が一番薄い
ハザール人やタタール人は長い間征服者だったのだ。カルムイクのように東洋人顔を残したければ
イラン系やスラブ系との結婚は避けねばならない。形質は黒人が一番強く白人がその次で、黄人は一番弱い。
でも立場的には黒人が一番弱かった。最近は芸能・スポーツのトップや閣僚になる人も増えてきたが、せいぜい70年代以後の話。
実際は混血は白黄でも黒黄でも、純粋な人種よりモンゴロイド(ーOー)的な薄さが加わりマイルドな風味になるがな。東亜系=顔が一番薄い
>>428
ベラルーシはロシアよりリトアニアやポーランドの影響が強い
ベラルーシはロシアよりリトアニアやポーランドの影響が強い
大正三美人の写真見たけど
あの頃まではまだノッペリ顔を良しとする美的感覚が生きてたんだな
Wikipedia先生には関東大震災後あたりから日本の美人感覚が変わり始めたというようなことを書いてあるけれど
ハリウッドの進出とかが影響しているんだろうか
あの頃まではまだノッペリ顔を良しとする美的感覚が生きてたんだな
Wikipedia先生には関東大震災後あたりから日本の美人感覚が変わり始めたというようなことを書いてあるけれど
ハリウッドの進出とかが影響しているんだろうか
脳の機能に注目してはどうだろう。
端正な顔立ち(の美形)という表現があるが、ぶっちゃけ平均値ってことでしょ。
いろんなブサイクを集めてもCGで合成(平均的な顔)を作ると端正な顔立ちになる。
脳は視覚情報から無意識に理想的(端正)な容姿を作って美醜を判断するとか。
現代において現実の人間よりテレビや映画の中の人間を凝視するようになれば、当然美醜の基準も修正をうける。さらに劇中の役回りの刷り込みですよ。
テレビによく出る人が美人に見える(akbがだんだん可愛く見える)のと同じ効果があるのでは?
テレビ・映画の出演者、彫刻・絵画のモチーフがすべて黒人ならどうなると思う?
端正な顔立ち(の美形)という表現があるが、ぶっちゃけ平均値ってことでしょ。
いろんなブサイクを集めてもCGで合成(平均的な顔)を作ると端正な顔立ちになる。
脳は視覚情報から無意識に理想的(端正)な容姿を作って美醜を判断するとか。
現代において現実の人間よりテレビや映画の中の人間を凝視するようになれば、当然美醜の基準も修正をうける。さらに劇中の役回りの刷り込みですよ。
テレビによく出る人が美人に見える(akbがだんだん可愛く見える)のと同じ効果があるのでは?
テレビ・映画の出演者、彫刻・絵画のモチーフがすべて黒人ならどうなると思う?
438 :世界@名無史さん:2012/11/25(日) 20:38:25.96 0
http://junko717.exblog.jp/
I wanna be a pop star 君をもっと夢中にさせてあげるからね
テカテカの小便小僧 羽根を広げ小便小僧にしてあげよう君だけに
君に出会えた喜びと君に会えない淋しさの
両方を手に入れて小便小僧は走り出す
空も飛べない僕だけど孤独を謳う夜だけど
その頬に 微笑みを与えられたなら
初めて君を抱きしめた瞬間に
神様が僕に下した使命は君だけの小便小僧
I wanna be a pop star 君をもっと夢中にさせてあげるからね
キラキラのpop star 羽根を広げ小便小僧をにしてあげよう
I wanna be your pop star 君をぎゅっと小便小僧にしてあげるからおいで
キラキラのpop star 羽根を広げ小便小僧にしてあげよう君だけに
隠していたい暗闇も君はそっと拾い上げて
大切な僕なんだとてくれた
光へと続く道を歩いて行こう
傷かばうこの右手は君と手をつなぐために
You're gonna be my pop star 僕をもっと小便小僧にさせてよ微笑んで
キラキラのpop star その瞳で僕を小便小僧に魔法をかけて
You are my only pop star 僕をぎゅっと抱きしめたらもう離さない
キラキラのpop star その瞳で僕を小便小僧に魔法をかけて今すぐに
恋に落ちたら誰もが誰かの小便小僧
神様が僕に下した使命は君だけの小便小僧
I wanna be a pop star 君をもっと夢中にさせてあげるからね
テカテカのpop star 羽根を広げ小便小僧に魔法をかけてあげよう
I wanna be your pop star 君をぎゅっと抱きしめて小便小僧にぃてあげるからおいで
キラキラのpop star 羽根を広げ小便小僧に魔法をかけてあげよう君だけに
I wanna be a pop star 君をもっと夢中にさせてあげるからね
テカテカの小便小僧 羽根を広げ小便小僧にしてあげよう君だけに
君に出会えた喜びと君に会えない淋しさの
両方を手に入れて小便小僧は走り出す
空も飛べない僕だけど孤独を謳う夜だけど
その頬に 微笑みを与えられたなら
初めて君を抱きしめた瞬間に
神様が僕に下した使命は君だけの小便小僧
I wanna be a pop star 君をもっと夢中にさせてあげるからね
キラキラのpop star 羽根を広げ小便小僧をにしてあげよう
I wanna be your pop star 君をぎゅっと小便小僧にしてあげるからおいで
キラキラのpop star 羽根を広げ小便小僧にしてあげよう君だけに
隠していたい暗闇も君はそっと拾い上げて
大切な僕なんだとてくれた
光へと続く道を歩いて行こう
傷かばうこの右手は君と手をつなぐために
You're gonna be my pop star 僕をもっと小便小僧にさせてよ微笑んで
キラキラのpop star その瞳で僕を小便小僧に魔法をかけて
You are my only pop star 僕をぎゅっと抱きしめたらもう離さない
キラキラのpop star その瞳で僕を小便小僧に魔法をかけて今すぐに
恋に落ちたら誰もが誰かの小便小僧
神様が僕に下した使命は君だけの小便小僧
I wanna be a pop star 君をもっと夢中にさせてあげるからね
テカテカのpop star 羽根を広げ小便小僧に魔法をかけてあげよう
I wanna be your pop star 君をぎゅっと抱きしめて小便小僧にぃてあげるからおいで
キラキラのpop star 羽根を広げ小便小僧に魔法をかけてあげよう君だけに
>>433
宗教ではないだろうか。
トルコ系はイスラム化したからコーカソイドに。
言いたいことはわかるが。対等混血の場合、白人的形質の方が強い、と言いたいのだろうけど。
でも北東アジアののっぺり顔が数千年で世界15億に増えたのも、
十分強い形質だと思う。
争う内容でもないが、
日本人から見てハーフは白人顔だが、
西欧から見てハーフは東洋人顔に見えるらしいので、
互いに相手の形質を多く見えるものではないだろうか。
宗教ではないだろうか。
トルコ系はイスラム化したからコーカソイドに。
言いたいことはわかるが。対等混血の場合、白人的形質の方が強い、と言いたいのだろうけど。
でも北東アジアののっぺり顔が数千年で世界15億に増えたのも、
十分強い形質だと思う。
争う内容でもないが、
日本人から見てハーフは白人顔だが、
西欧から見てハーフは東洋人顔に見えるらしいので、
互いに相手の形質を多く見えるものではないだろうか。
レーニンはともかくスターリン以降のソ連指導者は日本人から見れば白人だが。
ロシアやウクライナでコサックと呼ばれるのはスラブ系ではあるがトルコ系風俗を真似た人
国境地帯でトルコ系と戦ううち騎馬戦闘を取り入れた
国境地帯でトルコ系と戦ううち騎馬戦闘を取り入れた
プーチンなんか案外近所のおっさんにいそうではないか。
一般的にコサックはスラブ没落貴族等と言われているが、
本当は改宗トルコ系じゃないのだろうか。
コサックってウクライナ等では誇りの象徴だから、
スラブとしているが、西国からみた武士みたいなもので
発端は異民族なんじゃないの。
ウクライナとロシアはお互いを東方混血と言い罵り合ってるが、
両方とも矛盾してるな。
本当は改宗トルコ系じゃないのだろうか。
コサックってウクライナ等では誇りの象徴だから、
スラブとしているが、西国からみた武士みたいなもので
発端は異民族なんじゃないの。
ウクライナとロシアはお互いを東方混血と言い罵り合ってるが、
両方とも矛盾してるな。
武士が異民族?
自称アーリア人ヒムラーがどうしても
日本のサラリーマンの課長クラスに見えてしまう件について。
日本のサラリーマンの課長クラスに見えてしまう件について。
446 :世界@名無史さん:2013/01/02(水) 15:10:12.38 0
そこは論点じゃないw
そのように言われているだけで、
実際はスラブではない、という話。
そのように言われているだけで、
実際はスラブではない、という話。
448 :世界@名無史さん:2013/01/02(水) 16:40:50.39 0
>>447
スラブが人種や血統じゃなくて語派に過ぎないんだからその論点のほうがよっぽどおかしいよ
そもそもヤムナ文化など先史時代文化の時代からウクライナというのはいろんな人々がやってきて混ざっていた土地
そういう地理的な特徴がある
ウクライナ人はトルコ系というよりもタタールやゴートや遊牧スキタイやサルマタイなどの血が濃い
そして言語的にはスラブで、文化的にもあのウクライナの地域に独特の伝統を持つスラブ文化
中世前期のアント人というのもそういった流れの人々
スラブが人種や血統じゃなくて語派に過ぎないんだからその論点のほうがよっぽどおかしいよ
そもそもヤムナ文化など先史時代文化の時代からウクライナというのはいろんな人々がやってきて混ざっていた土地
そういう地理的な特徴がある
ウクライナ人はトルコ系というよりもタタールやゴートや遊牧スキタイやサルマタイなどの血が濃い
そして言語的にはスラブで、文化的にもあのウクライナの地域に独特の伝統を持つスラブ文化
中世前期のアント人というのもそういった流れの人々
449 :世界@名無史さん:2013/01/02(水) 16:45:19.39 0
>>447
コサックは右岸ウクライナの逃亡農民で、
土地の地主貴族たちが彼らの先祖にお金を貸して返せなくなったのを召し取って領地を広げていった
この傾向は16世紀後半に強まってコサックが増えた
そこでこの農民たちは農奴化されるのを嫌って東方のシーチに逃亡、自治を行いコサックとなった
彼らは農業に懲りたから農業はあまりやらず、もっぱら遊牧と、季節ごとにトルコ領(主にクリミア)へ略奪に行くことを生業としていた
シーチには各地からそういったアウトローたちが集まってきたから血統もバラバラで、
互いの意思を伝えるのに、仲間内で優勢だった当時のウクライナ訛りの東スラブ語(ルテニア語)をさらに訛って使っていた
そんなところ
コサックは右岸ウクライナの逃亡農民で、
土地の地主貴族たちが彼らの先祖にお金を貸して返せなくなったのを召し取って領地を広げていった
この傾向は16世紀後半に強まってコサックが増えた
そこでこの農民たちは農奴化されるのを嫌って東方のシーチに逃亡、自治を行いコサックとなった
彼らは農業に懲りたから農業はあまりやらず、もっぱら遊牧と、季節ごとにトルコ領(主にクリミア)へ略奪に行くことを生業としていた
シーチには各地からそういったアウトローたちが集まってきたから血統もバラバラで、
互いの意思を伝えるのに、仲間内で優勢だった当時のウクライナ訛りの東スラブ語(ルテニア語)をさらに訛って使っていた
そんなところ
450 :世界@名無史さん:2013/01/02(水) 16:49:29.43 0
もちろん彼らの女の調達はクリミアなど主にトルコ領、ついでフィン系諸部族のいる北方から
だから血統的にはごっちゃごちゃ
この連中がそれまでの支配層だったポーランド国家のウクライナ地方の
地主貴族たち(キエフ・ルーシ系からリトアニア大公国まで続いたルテニア人シュラフタ)にかわって
17世紀以降(特にボグダン・フメリニツキーの乱以降)にウクライナを支配していった
だから血統的にはごっちゃごちゃ
この連中がそれまでの支配層だったポーランド国家のウクライナ地方の
地主貴族たち(キエフ・ルーシ系からリトアニア大公国まで続いたルテニア人シュラフタ)にかわって
17世紀以降(特にボグダン・フメリニツキーの乱以降)にウクライナを支配していった
451 :世界@名無史さん:2013/01/02(水) 16:50:51.08 0
だからね、いまのウクライナがキエフ・ルーシの系統を強調しその正当性を主張するのは、どこか片腹痛い思いがする
コサックの文化はスキタイとかフンとか大ブルガール、ハザールに近い
そしてスラブ語派の地方文化の一つでもある
それがあのウクライナという土地の伝統的な歴史的発展形態
ウクライナとは限らないけれども、
スキタイもフンもブルガールもハザールもみな部族としてはスラブ語派の諸部族に取り込まれて消えていったね
それがあのウクライナという土地の伝統的な歴史的発展形態
ウクライナとは限らないけれども、
スキタイもフンもブルガールもハザールもみな部族としてはスラブ語派の諸部族に取り込まれて消えていったね
シーチは印欧語で城塞(の街)を意味する
フランス語のシテ、英語のシティーと同じ
ツィタデルなどの言葉も同語源
フランス語のシテ、英語のシティーと同じ
ツィタデルなどの言葉も同語源
>>448
中心をどちらと定義しているかの話。
ウクライナに遊牧系が入っているのではなく、
ウクライナそのものが遊牧系の系譜を含むのが実体。
だが解釈上は、スラブ・正教徒中心の歴史とされていることへの指摘。
中心をどちらと定義しているかの話。
ウクライナに遊牧系が入っているのではなく、
ウクライナそのものが遊牧系の系譜を含むのが実体。
だが解釈上は、スラブ・正教徒中心の歴史とされていることへの指摘。
なぜスラブが中心なのかといえばスラブ言語だから
いくら遊牧民が入ってきても最終的に当地のスラブ語派の諸部族の文化に吸収されて消えてしまう
そんな柔軟性があるのがウクライナにおけるスラブ語派の諸部族の伝統文化だった
外来の遊牧民がどんなに王を自称しても、結局消えてしまうのはその外来種の自称支配層だった。
政治的支配層(主に遊牧民たち)は文化的支配層(土着のスラブ系)とは異なる概念、と言ったほうがわかりやすいか。
いくら遊牧民が入ってきても最終的に当地のスラブ語派の諸部族の文化に吸収されて消えてしまう
そんな柔軟性があるのがウクライナにおけるスラブ語派の諸部族の伝統文化だった
外来の遊牧民がどんなに王を自称しても、結局消えてしまうのはその外来種の自称支配層だった。
政治的支配層(主に遊牧民たち)は文化的支配層(土着のスラブ系)とは異なる概念、と言ったほうがわかりやすいか。
おそらくだが、ウクライナのスラブ人たちがそれほどまでに強い理由は、
彼らが古くから農耕をしっかり行っていたからだろうね
彼らの伝統である穀物生産は、外来の種族同士の政治的争いに関係なく行われるから、
政権が代わっても敗者たちがスラブ系土着民の社会に混じってしまって、
独立した集団としては消えてしまう
そんな集団は歴史に記述されるだけでも、
スキタイ、サルマタイ、ゴート、フン、アヴァール、ブルガール、ハザール、マジャール、モンゴル、など
彼らが古くから農耕をしっかり行っていたからだろうね
彼らの伝統である穀物生産は、外来の種族同士の政治的争いに関係なく行われるから、
政権が代わっても敗者たちがスラブ系土着民の社会に混じってしまって、
独立した集団としては消えてしまう
そんな集団は歴史に記述されるだけでも、
スキタイ、サルマタイ、ゴート、フン、アヴァール、ブルガール、ハザール、マジャール、モンゴル、など
その定義であるならコサックは改宗トルコ人で問題ないのでは。
武士はアズマエビスと言う明らかな異民族だよ。
ただそんな発想は口に出さないだけで。
武士はアズマエビスと言う明らかな異民族だよ。
ただそんな発想は口に出さないだけで。
459 :世界@名無史さん:2013/01/02(水) 23:48:28.75 0
>>458
ないない。
おまえ血統と言語集団を混同しているよ。
〜人というときはまずは言語集団のことなのが常識。
血統と言語の分布が一致しないときは言語を取り、血統は取らない。
その場合…系〜人という言い方はするだろう。だがそれでも〜人である。
じゃウクライナ人は言語を取ってスラヴ人だとして何系なのか?
ウクライナは歴史時代に入っても多種の混血が行われた土地だから、
特定の一つの先祖を取ってそれ系だとは言えない。
例えばウクライナ人に対してトルコ系スラヴ人という呼び方が正しいか?答えは否。
ないない。
おまえ血統と言語集団を混同しているよ。
〜人というときはまずは言語集団のことなのが常識。
血統と言語の分布が一致しないときは言語を取り、血統は取らない。
その場合…系〜人という言い方はするだろう。だがそれでも〜人である。
じゃウクライナ人は言語を取ってスラヴ人だとして何系なのか?
ウクライナは歴史時代に入っても多種の混血が行われた土地だから、
特定の一つの先祖を取ってそれ系だとは言えない。
例えばウクライナ人に対してトルコ系スラヴ人という呼び方が正しいか?答えは否。
461 :世界@名無史さん:2013/01/03(木) 01:14:44.57 0
なんで中世コサック(14世紀)の話になるんだ?
そもそも、すくなくともその時代からずっと文献に出てくるコサックはみなルテニア語話してるだろ。
それじゃ不十分なのか?
またチュルク系を母語にしていたのはタタール人だ、混同すんな。
そもそも、すくなくともその時代からずっと文献に出てくるコサックはみなルテニア語話してるだろ。
それじゃ不十分なのか?
またチュルク系を母語にしていたのはタタール人だ、混同すんな。
現代にコサックなんていねえよ。
日本に武士探しに来る外人かお前は。
コサックという単語がテュルクなのに。
タタールはモンゴルのことだつうの。
今のタタール語だってもうほとんど話されてないのにタタール人と呼ぶだろ。
日本に武士探しに来る外人かお前は。
コサックという単語がテュルクなのに。
タタールはモンゴルのことだつうの。
今のタタール語だってもうほとんど話されてないのにタタール人と呼ぶだろ。
463 :世界@名無史さん:2013/01/03(木) 01:57:37.57 0
中世じゃないと現代なのかおめでたいな。
コサックが最も勢力を持っていた近代が抜けてるぞ。
しかもコサックがスラヴでないなんて屁理屈誰も認めないよ。
コサックが最も勢力を持っていた近代が抜けてるぞ。
しかもコサックがスラヴでないなんて屁理屈誰も認めないよ。
お前だけだろ。
タタールも知らんくせに。黙って年号でも暗記してろ。
タタールも知らんくせに。黙って年号でも暗記してろ。
465 :世界@名無史さん:2013/01/03(木) 12:14:45.73 0
コサックをトルコ人と呼ぶなら、ドイツ人をスラブ人と呼ばなくてはならない。
そしてロシア人をスラブ人でなくフィン人と呼ぶべきだろう。ポーランド人はゲルマン人と呼んだ方がいい。
そしてロシア人をスラブ人でなくフィン人と呼ぶべきだろう。ポーランド人はゲルマン人と呼んだ方がいい。
呼べば。
コサックは民族ではないし。
ロシアが束ねていた同盟軍のひとつだよ。
コサックは民族ではないし。
ロシアが束ねていた同盟軍のひとつだよ。
たしかに外人(東アジア系)は醜いよな
471 :世界@名無史さん:2013/03/24(日) 12:42:17.28 0
>>468
1月3日の昼間の書き込みに対してニートと決めつけるということは。。^^;
1月3日の昼間の書き込みに対してニートと決めつけるということは。。^^;
ドイツで一時期暮らしてたけどブスしかいなかったけどな。
その上、ゴリラみたいなゴツさで近づくと髭が生えてるから手に負えない。
たまに可愛いの見つけると東欧系の移民だったりさ。
イギリスも似た感じだった。
その上、ゴリラみたいなゴツさで近づくと髭が生えてるから手に負えない。
たまに可愛いの見つけると東欧系の移民だったりさ。
イギリスも似た感じだった。
473 :世界@名無史さん:2013/04/14(日) 18:29:15.14 I BE:2027635564-2BP(0)
戦争で一度も勝った事のない小国島国が大日本帝国だなんて笑わせる
受け狙いで名付けたのか?
受け狙いで名付けたのか?
大韓民国様乙
475 :世界@名無史さん:2013/05/22(水) 00:40:44.66 0
こういうの見つけた。>>10 は俺の投稿だが図を貼った覚えが無いし、
このスレの >>10 にも無い。どうなってるだろ?
http://blog.livedoor.jp/gurigurimawasu/archives/19177761.html
このスレの >>10 にも無い。どうなってるだろ?
http://blog.livedoor.jp/gurigurimawasu/archives/19177761.html
>>472
そう。白人綺麗と言うが、
アメリカ映画観てても不細工役がいっぱい出てくるワケで。
美人の方がブスより多いのは中東、ロシアだけかと。
で、
その地域に対しては、コンプレックス無いよな。
ロシアは二等白人で、中東は賢く無い。
欧米コンプレックスがあり、且つ容姿淡麗で参りましたと言えるのは
フランスと、イタリア北部と、北欧と、アイルランドかな?
ドイツ、イギリスは綺麗と汚いに差がありすぎ。
そう。白人綺麗と言うが、
アメリカ映画観てても不細工役がいっぱい出てくるワケで。
美人の方がブスより多いのは中東、ロシアだけかと。
で、
その地域に対しては、コンプレックス無いよな。
ロシアは二等白人で、中東は賢く無い。
欧米コンプレックスがあり、且つ容姿淡麗で参りましたと言えるのは
フランスと、イタリア北部と、北欧と、アイルランドかな?
ドイツ、イギリスは綺麗と汚いに差がありすぎ。
477 :世界@名無史さん:2013/05/22(水) 04:11:55.65 0
なんで白人は美しいかを考察した本をここに紹介しろ。
俺様が読んでやるから。
俺様が読んでやるから。
478 :世界@名無史さん:2013/05/23(木) 00:09:59.84 O
>ロシアは二等白人で
それでも世界初の宇宙飛行
それでも世界初の宇宙飛行
ヨーロッパ人といっても多様性があるわけで
地中海系やスラブ系はアジア系より可愛いのが多いと思うけど
現在正統白人とされてるコテコテのゲルマン系の女は>>472が酷評するように
俺の中では黒人女と同じ最下層カーストだわ
地中海系やスラブ系はアジア系より可愛いのが多いと思うけど
現在正統白人とされてるコテコテのゲルマン系の女は>>472が酷評するように
俺の中では黒人女と同じ最下層カーストだわ
アフガニスタンが容姿端麗。
中東とか顔濃いから美しいはならんだろ
中国に昔イラン人居ても美しいにはならなかったしな
やはりあまり濃くないロシアとかエゲレスが美しくなるわけね
中国に昔イラン人居ても美しいにはならなかったしな
やはりあまり濃くないロシアとかエゲレスが美しくなるわけね
濃いのはアラブ。
イギリスは無い。
イギリスは無い。
ロシアはともかくイギリスってw
まあ美的感覚は個人個人だし・・・・
俺的にはロシア>>イギリス>>ドイツだが
俺的にはロシア>>イギリス>>ドイツだが
ロシアじゃなくてウクライナね。
これはロシア人も認めとる。ロシアにはハズレも多いがウクライナは少ないけぇのう
これはロシア人も認めとる。ロシアにはハズレも多いがウクライナは少ないけぇのう
朝鮮人が必死に日本人は白人コンプとか連呼してるけど
そういう連呼してるのが白人コンプ臭い
そういう連呼してるのが白人コンプ臭い
淡麗生。
イギリスドイツオランダ
この辺の女はゴツゴツしてそうなイメージ
同系統の北欧系はもう少し柔らかくした印象なんだが
オランダ人の男は自国の女性より小柄で女性らしさがあるイタリア、スペイン系のほうが好きらしい
この辺の女はゴツゴツしてそうなイメージ
同系統の北欧系はもう少し柔らかくした印象なんだが
オランダ人の男は自国の女性より小柄で女性らしさがあるイタリア、スペイン系のほうが好きらしい
490 :世界@名無史さん:2013/07/04(木) 14:00:31.26 ID:drMAFzh+0!
オランダ人の両親の下で生まれ育ったサッカーのハーフナーもこの前のインタビューで「オランダ人はでかすぎ。日本の女性のほうが良い」とか言ってたね
ドイツ在住だが、マジでこっちの女はごつい
行く前はオクトーバーフェストの画像とか見てwkwkしてたんだが
仕込みとコスチューム補正が働いてたんだなと気づかされ酷い目に遭った
行く前はオクトーバーフェストの画像とか見てwkwkしてたんだが
仕込みとコスチューム補正が働いてたんだなと気づかされ酷い目に遭った
492 :世界@名無史さん:2013/07/04(木) 18:01:44.16 0
うちのジュモーちゃんも白人のはず
Missドイツ候補
http://img.thesun.co.uk/multimedia/archive/00426/germanymain_426245a.jpg
オールチェンジ
修学旅行でバスの中で盛り上がるドイツの女子高生達
http://livedoor.blogimg.jp/new_sappukei/imgs/7/e/7e60caaf-s.jpg
http://livedoor.blogimg.jp/new_sappukei/imgs/f/e/fe7491e8-s.jpg
http://livedoor.blogimg.jp/new_sappukei/imgs/6/b/6ba5b4da-s.jpg
喧嘩したら負けそうなんですけど・・・
http://img.thesun.co.uk/multimedia/archive/00426/germanymain_426245a.jpg
{kind=link}
オールチェンジ
修学旅行でバスの中で盛り上がるドイツの女子高生達
http://livedoor.blogimg.jp/new_sappukei/imgs/7/e/7e60caaf-s.jpg
{kind=link}
http://livedoor.blogimg.jp/new_sappukei/imgs/f/e/fe7491e8-s.jpg
{kind=link}
http://livedoor.blogimg.jp/new_sappukei/imgs/6/b/6ba5b4da-s.jpg
{kind=link}
喧嘩したら負けそうなんですけど・・・
フェティシズム大国ドイツ(SM、スカトロなど)
>>493
た、たくましい・・・・
た、たくましい・・・・
ジュモーってアンティークドールかよwww
ラテン系だろ?
ラテン系だろ?
497 :世界@名無史さん:2013/07/06(土) 13:29:13.44 0
>>493
ヒトラーが「ユダヤ人が我々の美しいドイツ人女性と結婚して血を汚してる」とか言ってたけど自意識過剰!!
韓国人が「ウリ達は美男子ニダ」と言ってるのと同じくらい勘違いで滑稽。
イスラエルのアシュケナージ系ユダヤ人の女性兵士は美人で頭も良くて強くて。
http://blog-imgs-11.fc2.com/t/o/m/tomoyan3/20070320143144.jpg
ヒトラーが「ユダヤ人が我々の美しいドイツ人女性と結婚して血を汚してる」とか言ってたけど自意識過剰!!
韓国人が「ウリ達は美男子ニダ」と言ってるのと同じくらい勘違いで滑稽。
イスラエルのアシュケナージ系ユダヤ人の女性兵士は美人で頭も良くて強くて。
http://blog-imgs-11.fc2.com/t/o/m/tomoyan3/20070320143144.jpg
{kind=link}
498 :世界@名無史さん:2013/07/06(土) 13:51:57.25 0
白人(ゲルマン)コンプレックス総合 32コンプ目
http://kohada.2ch.net/test/read.cgi/diet/1367313182/l50
http://kohada.2ch.net/test/read.cgi/diet/1367313182/l50
まぁつい2000年くらい前まではゲルマン人は森でウホウホやってた黒人と大差ない連中だったからブスばっかで当然
500 :世界@名無史さん:2013/07/06(土) 14:27:38.16 0
500get
ビッチな欧米女より、イラン女性のほうが好きです!!
502 :世界@名無史さん:2013/07/12(金) 05:33:35.28 0
>>59
そもそも洋服自体が日本のものではないからな
そもそも洋服自体が日本のものではないからな
503 :世界@名無史さん:2013/07/12(金) 05:42:16.89 0
性選択がおおいに影響していると思いますが、いかがでしょうか?
ラテン人が現代ヨーロッパ白人の基となるケルト人、ゲルマン人を占領地でレイプしまくって
そしてノルマン人もスラブ人を含めた西欧東欧南欧各地でレイプしまくった。
東アジアで言えば弥生系の古代漢民族がポリネシア人や東南アジア人を征服してレイプしまくり
さらにアイヌや縄文系、チベット人などとも混血しまくってるようなもの。
血が混ざりまくったから美人が多いのだろ
そしてノルマン人もスラブ人を含めた西欧東欧南欧各地でレイプしまくった。
東アジアで言えば弥生系の古代漢民族がポリネシア人や東南アジア人を征服してレイプしまくり
さらにアイヌや縄文系、チベット人などとも混血しまくってるようなもの。
血が混ざりまくったから美人が多いのだろ
血が混ざらなかったら不細工になるの?
アボリジニとか?
アボリジニとか?
韓国人とか
韓国人って純潔なの?
結構侵略とかで異民族の侵入が多いと思うけど。
結構侵略とかで異民族の侵入が多いと思うけど。
侵略されてもブスばっかで言われてるほどレイプされてないんじゃないかな
それよりも歴代中華の犯罪者や逃亡奴隷の掃き溜めで美人が出ても中華やモンゴルに貢いで
近親相姦ばっかしてたからあんな不細工な上に歪んだ民族性になっちゃったんじゃないか
それよりも歴代中華の犯罪者や逃亡奴隷の掃き溜めで美人が出ても中華やモンゴルに貢いで
近親相姦ばっかしてたからあんな不細工な上に歪んだ民族性になっちゃったんじゃないか
半島への犯罪者や奴隷の逃亡ってあったの?
船を調達するか女真族の土地を突破しなきゃならなくて面倒だと思うけど
船を調達するか女真族の土地を突破しなきゃならなくて面倒だと思うけど
511 :世界@名無史さん:2013/09/02(月) 05:07:07.48 0
こういう頭の悪そうなスレはネトウヨが湧くのね
512 :世界@名無史さん:2013/09/02(月) 10:52:42.14 0
美の基準が西洋だからなねえ
だから西洋人に近い中東とかインドとかも容姿はいい
だから西洋人に近い中東とかインドとかも容姿はいい
欧米はごつすぎる
スラヴは可愛いしイランは麗しい
スラヴは可愛いしイランは麗しい
514 :世界@名無史さん:2013/09/28(土) 00:13:47.98 0
ラテン系はアラブ人の血が混じってる。
というかアラブ人でキリスト教化したのがラテン系。
だから浅黒くくて天パが多いんだよ。
というかアラブ人でキリスト教化したのがラテン系。
だから浅黒くくて天パが多いんだよ。
515 :世界@名無史さん:2013/09/28(土) 08:46:06.47 0
明治政府はナポレオン生誕100周年記念日に発足
http://ichiba.geocities.jp/gbsg0309/0203/35/237.html
これは、フランス人が日本を支配したのではなく、
ヨーロッパは、最初から天皇一族の支配地だったことを意味する。
(仮説を含む)
( http://ichiba.geocities.jp/gbsg0309/0103/34/231.html )
516 :(スリーサイズスリーサイズスリーサイズスリーサイズフリーサイズ):2013/09/28(土) 15:20:46.67 0
>ヨーロッパ人がなんで容姿淡麗か
政経
政経
東アジアはのっぺりしすぎ
のっぺらぼうは朝鮮だけ
中国人は目元は普通だが鼻が高い
モンゴル人は丸顔だが身体能力が高い
朝鮮だけが容貌醜悪で基礎体力なし頭もおかしい
中国人は目元は普通だが鼻が高い
モンゴル人は丸顔だが身体能力が高い
朝鮮だけが容貌醜悪で基礎体力なし頭もおかしい
むなしいね〜
アメリカの黒人がアフリカの黒人に「あいつらの方が黒い」とか言ってるようなもの
アメリカの黒人がアフリカの黒人に「あいつらの方が黒い」とか言ってるようなもの
人って肌の色よりも顔のパーツの方に目がいくと思うんだがどうよ
M・ジャクソンも黒人特有の幅広の鼻を嫌って何度も整形でいじくって
仕舞いには鼻自体がもげちゃったって話しだし
M・ジャクソンも黒人特有の幅広の鼻を嫌って何度も整形でいじくって
仕舞いには鼻自体がもげちゃったって話しだし
>>520
11世紀イランの『カーブースの書』に載っている奴隷購入についての助言によると
「インド人は顔の一つ一つの部分を見ると美しいが、顔全体としてみると美しくない。
トルコ人(この頃は突厥などの後裔のアジア系遊牧民を指す)は
顔の一つ一つの部分は美しくないが、顔全体として見ると美しい」
11世紀イランの『カーブースの書』に載っている奴隷購入についての助言によると
「インド人は顔の一つ一つの部分を見ると美しいが、顔全体としてみると美しくない。
トルコ人(この頃は突厥などの後裔のアジア系遊牧民を指す)は
顔の一つ一つの部分は美しくないが、顔全体として見ると美しい」
522 :フランス語でロリジュンって言うんだぜ:2013/10/03(木) 19:33:33.62 0
フランスだと歯が生えても赤ちゃんにおしゃぶりさせてるって聞いたことがあるけど、どうなん?
523 :世界@名無史さん:2013/10/04(金) 01:19:45.40 O
一般人は別に容姿端麗でも何でもない
確かに町で見かける子供たちはニグロや蒙古の子供とは全く別次元の可愛いらしさがある
だが成長期を過ぎると急速にネアン化が進む
だが成長期を過ぎると急速にネアン化が進む
525 :世界@名無史さん:2013/10/04(金) 06:06:13.39 O
日本人がみた白人の美人と
白人がみた白人の美人は違うだろ
日本人がみた日本人の美人と
白人がみた日本人の美人も違う
特に黒人兵士が連れてる日本人女って鼻が低くて顔の真ん中潰れたような女が多いだろ
アメリカ映画のヒロインも 顎が片桐はいりみたいな女が多い
日本人好みの白人女はたいてい脇役で どこと無く日本人的要素がまじった女が多い
白人がみた白人の美人は違うだろ
日本人がみた日本人の美人と
白人がみた日本人の美人も違う
特に黒人兵士が連れてる日本人女って鼻が低くて顔の真ん中潰れたような女が多いだろ
アメリカ映画のヒロインも 顎が片桐はいりみたいな女が多い
日本人好みの白人女はたいてい脇役で どこと無く日本人的要素がまじった女が多い
美の価値観にそれほど差があるとも思えんが
東アジア人の特徴はコーカソイドとは真逆なのは致命的だな
コーカソと蒙古の発生地は距離的に極端に離れていないんだけどね
不思議なもんだ
不思議なもんだ
>>526
差はあるよ。纏足を綺麗だと思うか?リングで長くした首を綺麗だと思うか?
差はあるよ。纏足を綺麗だと思うか?リングで長くした首を綺麗だと思うか?
530 :カモッラって闇パン売ってるの知ってる?:2013/10/04(金) 21:05:48.54 0
東スラヴはのっぺりしすぎ
19世紀末から20世紀初頭にかけて朝鮮半島に訪れた白人は
朝鮮人の容姿を誉めまくってるよね
朝鮮人の容姿を誉めまくってるよね
*ばっちいので触らないように
いやほんとなんだよ
勿論不潔さも非難してたけどさ
あれは白人にとって体格の立派さこそが重要だったのか
それとも本当に白人の美的感覚では朝鮮人が美形に見えたのか
前知識から認識にバイアスがかかっていただけなのか
どう解釈したらいいのかよく分からなくてね
ここの人の意見を聞きたい
勿論不潔さも非難してたけどさ
あれは白人にとって体格の立派さこそが重要だったのか
それとも本当に白人の美的感覚では朝鮮人が美形に見えたのか
前知識から認識にバイアスがかかっていただけなのか
どう解釈したらいいのかよく分からなくてね
ここの人の意見を聞きたい
535 :世界@名無史さん:2013/10/05(土) 06:20:51.57 O
エキゾチシズムじゃね
あと未開国なのにってバイアス
日本人と違い 肉食してたから体格も上だろう
ちなみにかつて世界一の美人国はタイだと聞いたことあるが
あと未開国なのにってバイアス
日本人と違い 肉食してたから体格も上だろう
ちなみにかつて世界一の美人国はタイだと聞いたことあるが
ニューハーフはタイが世界一
>>534
チョンのデマカセか最大限歪曲拡大した解釈かのどちらかだろ
少なくとも現存する写真に艶やかな姿形の人間なぞただの一人も写っとらん
むしろ同時代の写真に写る日本の大名とかの方がよほど文明人の顔つきだし
多分真っ黒で大きいニグロが美しいとかゆう類と同レベルの話
チョンのデマカセか最大限歪曲拡大した解釈かのどちらかだろ
少なくとも現存する写真に艶やかな姿形の人間なぞただの一人も写っとらん
むしろ同時代の写真に写る日本の大名とかの方がよほど文明人の顔つきだし
多分真っ黒で大きいニグロが美しいとかゆう類と同レベルの話
538 :世界@名無史さん:2013/10/06(日) 15:47:47.99 O
>537
ガラス玉の中の水晶と
石ころの中の水晶
どちらが目立つ?
ガラス玉の中の水晶と
石ころの中の水晶
どちらが目立つ?
539 :貨幣にしとけばいいものを:2013/10/06(日) 16:04:39.80 0
ああ、ダイヤってアフリカじゃ石ころ扱いなんだけ?
541 :世界@名無史さん:2013/10/08(火) 15:14:29.37 0
>>531 逆だろ?ギリシャ彫刻ににより近いのがスラブ系。
スラブより地理的にも血縁的にもモンゴルに遠いはずのゲルマン系の方が
地黒やのっぺり顔が多くね?歯が汚くて歯列矯正が必要な奴もスラブやラテンより多い。
あとスラブやゲルマンの金髪は地黒のラテン系より劣化しやすいよな。
ラテン・モンゴル女は五十にして劣化し、ゲルマン・スラブ女は十五にして劣化する。
美しかった妖精が!!!悲しいかな白人や黒人には日本みたいな合法ロリは出来んのだ。
スラブより地理的にも血縁的にもモンゴルに遠いはずのゲルマン系の方が
地黒やのっぺり顔が多くね?歯が汚くて歯列矯正が必要な奴もスラブやラテンより多い。
あとスラブやゲルマンの金髪は地黒のラテン系より劣化しやすいよな。
ラテン・モンゴル女は五十にして劣化し、ゲルマン・スラブ女は十五にして劣化する。
美しかった妖精が!!!悲しいかな白人や黒人には日本みたいな合法ロリは出来んのだ。
ゲルマンは角ばってるだろ。女も良く見ないと男に見えるくらい
東スラヴの方が丸っこくてノッペリしている。
東スラヴの方が丸っこくてノッペリしている。
見た目でゲルマン系とスラブ系の区別なんて出来るのか?スゲーな
スウェーデン人とサウジ人くらい違うとわかりやすいけど
イギリス人ドイツ人オランダ人とロシア人ポーランド人ウクライナ人なんて全然わからん
日本人と韓国人と中国人の違いが東アジア人以外にはわからないように
スウェーデン人とサウジ人くらい違うとわかりやすいけど
イギリス人ドイツ人オランダ人とロシア人ポーランド人ウクライナ人なんて全然わからん
日本人と韓国人と中国人の違いが東アジア人以外にはわからないように
>>537
気分次第によって、むやみに褒めたり、天気が悪く気分が良くないからと旅行した地区をぼろくそに貶す
イギリスの女性旅行作家にイザベラ・バード女史ってのがいる。
よく、2chのコピペにされる、朝鮮半島がいかに汚いのか、それで民衆が汚くても平気なのかのの文章を
書いた人だね。
彼女は、朝鮮半島の衛生環境が悪いことをこれでもか!って書いているけど、同時に朝鮮人の体格が良
く、容姿も優れていることも同時に表現しているよ。
公平を期すために両方目を通したら良いのでは?
ちなみに、国会図書館のデジタルライブラリーに「30年前の朝鮮」って書名で登録されているから
PCあったら誰でもただで読める。著者名は一時結婚してた名字に変わっているけどね。
気分次第によって、むやみに褒めたり、天気が悪く気分が良くないからと旅行した地区をぼろくそに貶す
イギリスの女性旅行作家にイザベラ・バード女史ってのがいる。
よく、2chのコピペにされる、朝鮮半島がいかに汚いのか、それで民衆が汚くても平気なのかのの文章を
書いた人だね。
彼女は、朝鮮半島の衛生環境が悪いことをこれでもか!って書いているけど、同時に朝鮮人の体格が良
く、容姿も優れていることも同時に表現しているよ。
公平を期すために両方目を通したら良いのでは?
ちなみに、国会図書館のデジタルライブラリーに「30年前の朝鮮」って書名で登録されているから
PCあったら誰でもただで読める。著者名は一時結婚してた名字に変わっているけどね。
俺が思うに典型的な新モンゴロイドの特質を持つ朝鮮人は顔は流線型でヒゲがほとんどない
体つきも筋肉質ではないので全体的にスラッとして見える
それが欧州人のイザベラ・バードにとって目新しく好意的な見方として記録されたのではないかな
いずれにせよイザベラ・バードの視点は内容の通り極めて俗物的でそれゆえ人種偏見を持つ人間にとっては第一級の資料となっている
体つきも筋肉質ではないので全体的にスラッとして見える
それが欧州人のイザベラ・バードにとって目新しく好意的な見方として記録されたのではないかな
いずれにせよイザベラ・バードの視点は内容の通り極めて俗物的でそれゆえ人種偏見を持つ人間にとっては第一級の資料となっている
欧州や中東は人種がコーカソイドだから彫りの深い顔立ちに高い鼻が標準装備
男の場合はカッコいいけど、女ならもう少しモンゴロイド的なのっぺり感があってもいい
男の場合はカッコいいけど、女ならもう少しモンゴロイド的なのっぺり感があってもいい
良い感じに過疎ってるのでGet!すます
548 :世界@名無史さん:2013/11/17(日) 10:49:42.14 0
とりあえず、これを元に話を始める
Ancient DNA Reveals Key Stages in the Formation of Central European Mitochondrial Genetic Diversity
http://www.sciencemag.org/content/342/6155/257.abstract
Ancient DNA Reveals Key Stages in the Formation of Central European Mitochondrial Genetic Diversity
http://www.sciencemag.org/content/342/6155/257.abstract
Early Neolithic 5500-3950 cal BC
Linear Pottery culture LBK 5500-4775 cal BC
Rossen culture RSC 4625-4250 cal BC
Schoningen group SCG 4100-3950 cal BC
Middle Neolithic 3950-2650 cal BC
Baalberge culture BAC 3950-3400 cal BC
Salzmunde culture SMC 3400-3100/3025 cal BC
Bernburg culture BEC 3100-2650 cal BC
Late Neolithic 2800-2200 cal BC
Corded Ware culture CWC 2800-2200/2050 cal BC
Bell Beaker culture BBC 2500-2200/2050 cal BC
Early Bronze Age 2200-1550 cal BC
Unetice culture UC 2200-1550 cal BC
Linear Pottery culture LBK 5500-4775 cal BC
Rossen culture RSC 4625-4250 cal BC
Schoningen group SCG 4100-3950 cal BC
Middle Neolithic 3950-2650 cal BC
Baalberge culture BAC 3950-3400 cal BC
Salzmunde culture SMC 3400-3100/3025 cal BC
Bernburg culture BEC 3100-2650 cal BC
Late Neolithic 2800-2200 cal BC
Corded Ware culture CWC 2800-2200/2050 cal BC
Bell Beaker culture BBC 2500-2200/2050 cal BC
Early Bronze Age 2200-1550 cal BC
Unetice culture UC 2200-1550 cal BC
ヨーロッパ、初期農耕時代に人口激減
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=20131002001
Dan Vergano
National Geographic News
October 2, 2013
ヨーロッパで農耕が始まって間もない時期には「饗宴か飢餓か」の極端な環境の変化が見られたことが、考古学の研究によって分かった。
放射性炭素年代測定法を用いた調査によると、フランスからアイルランドの初期農耕時代には、人口の爆発的増加の後に、極端な減少が確認できるという。
ヨーロッパにおける農耕は、約8500年前のギリシャにもたらされたのを端緒とし、それから数千年かけてアイルランドや北ヨーロッパにまで広がった。
狩猟採集から農耕への転換は人類史における大きな一歩ではあったが、当時の遺物からは、初期農耕時代の苛酷な状況がうかがえる。
「おそらく土壌の極度の荒廃が、疾病の流行や交戦につながったのだろう。当時の状況は気の重くなるようなも のだが、マルサスの(『人口論』での)主張のとおりでもある」と、
今回の論文の共著者の1人、ショーン・ダウニー(Sean Downey)氏は言う。ダウニー氏はメリーランド大学カレッジパーク校の考古学者だ。
ヨーロッパ最初の農耕民族は、森を切り開き種を蒔き穀物を育てながら、まずはフランス南部からデンマークまでの西ヨーロッパ一帯に勢力を拡大していったと、この論文では分析している。
研究チームは当時の土地利用と気候の状況を示すデータを、ヨーロッパ各地の考古学遺跡から確認された1万3658件の放射性炭素年代測定法による包括的データと対照した。
その結果、新石器時代の農耕経済は紀元前4000年頃にいったん衰退を迎えたことが推測された。
土地の利用状況や埋葬地の調査結果から、一部の地域ではこの時期に30〜60%も人口が減少していることが分かる。
中世ヨーロッパで“黒死病”(ペスト)が大流行した時期に匹敵する減少率である。「この人口の落ち込みは3〜6世紀にわたって続いた。単なる長期的な停滞期などというレベルではない」とダウニー氏は言う。
ヨーロッパの新石器時代においては、好条件の時期であっても農耕民族はそれほど人口を拡大できたわけではない。
現代のヨーロッパは、当時に比べると33倍も人口密度が高くなっている。
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=20131002001
Dan Vergano
National Geographic News
October 2, 2013
ヨーロッパで農耕が始まって間もない時期には「饗宴か飢餓か」の極端な環境の変化が見られたことが、考古学の研究によって分かった。
放射性炭素年代測定法を用いた調査によると、フランスからアイルランドの初期農耕時代には、人口の爆発的増加の後に、極端な減少が確認できるという。
ヨーロッパにおける農耕は、約8500年前のギリシャにもたらされたのを端緒とし、それから数千年かけてアイルランドや北ヨーロッパにまで広がった。
狩猟採集から農耕への転換は人類史における大きな一歩ではあったが、当時の遺物からは、初期農耕時代の苛酷な状況がうかがえる。
「おそらく土壌の極度の荒廃が、疾病の流行や交戦につながったのだろう。当時の状況は気の重くなるようなも のだが、マルサスの(『人口論』での)主張のとおりでもある」と、
今回の論文の共著者の1人、ショーン・ダウニー(Sean Downey)氏は言う。ダウニー氏はメリーランド大学カレッジパーク校の考古学者だ。
ヨーロッパ最初の農耕民族は、森を切り開き種を蒔き穀物を育てながら、まずはフランス南部からデンマークまでの西ヨーロッパ一帯に勢力を拡大していったと、この論文では分析している。
研究チームは当時の土地利用と気候の状況を示すデータを、ヨーロッパ各地の考古学遺跡から確認された1万3658件の放射性炭素年代測定法による包括的データと対照した。
その結果、新石器時代の農耕経済は紀元前4000年頃にいったん衰退を迎えたことが推測された。
土地の利用状況や埋葬地の調査結果から、一部の地域ではこの時期に30〜60%も人口が減少していることが分かる。
中世ヨーロッパで“黒死病”(ペスト)が大流行した時期に匹敵する減少率である。「この人口の落ち込みは3〜6世紀にわたって続いた。単なる長期的な停滞期などというレベルではない」とダウニー氏は言う。
ヨーロッパの新石器時代においては、好条件の時期であっても農耕民族はそれほど人口を拡大できたわけではない。
現代のヨーロッパは、当時に比べると33倍も人口密度が高くなっている。
>>550
この大幅な人口の落ち込みを経て、紀元前2800年頃には再び小規模な人口増加が見られる。ただし、人口の増加と減少のいずれも、気象条件との相関は見られなかった。
これは専門家にとって驚くべき結果だった。というのも、多くの場合、人口変動には気象が関係していると考えられているからだ。
たとえば3月に「Science」誌に発表された別の研究では、古典期マヤ文明が紀元800年頃に衰退した主な原因は干ばつであったと指摘されている。
「今回の研究結果を元に、今後いくつもの新たな研究が行われるものと確信している」と、スペイン、ジローナ大学のネウス・イサーン(Neus Isern)氏は言う。
イサーン氏は人口モデリングの専門家である。「背景に自然災害がなかったというのなら、新石器時代の経済はなぜ破綻したのか。新石器時代の経済は、私たちが考えるほど持続的なものではなかったのだろうか?」
ダウニー氏が推測しているのは、初期農耕民族は森林の乱伐や農地の継続使用によって急速な土壌の荒廃を招き、その結果、疾病が流行しやすくなって、人口減少の引き金を引いたというシナリオだ。
もう1つの可能性として、アイルランドにあるユニバーシティ・カレッジ・コークの考古学者ロン・ピンハシ(Ron Pinhasi)氏が提唱するのは、人口の大幅な増減の主な原因を移住であるとする考え方だ。
「2度目の人口増加は、乳製品のような二次生産物が作られはじめるという革新の結果であった可能性がある。酪農じたいは以前から始まっていたが、各地に広がるのは後になってからだ」とピンハシ氏は言う。
一般的にヒトの成人は牛乳などに含まれる乳糖を腸で分解できないが、ヨーロッパでは遺伝子変異のおかげで、90%近い成人が牛乳を飲める。
2009年の研究によると、この遺伝子変異はわずか7500年ほど前に起こった可能性が高いという。
「この研究結果には、現代一般的な考え方に対する、非常に重要なメッセージが含まれていると思わずにいられない。
現代では、新たなテクノロジーこそが常に時代を制するという盲信があちこちに見られるが、それは新石器時代には当てはまらないのだから」とダウニー氏は言う。
ヨーロッパの初期農耕時代に関する今回の研究は、「Nature Communications」誌に10月1日に掲載された。
この大幅な人口の落ち込みを経て、紀元前2800年頃には再び小規模な人口増加が見られる。ただし、人口の増加と減少のいずれも、気象条件との相関は見られなかった。
これは専門家にとって驚くべき結果だった。というのも、多くの場合、人口変動には気象が関係していると考えられているからだ。
たとえば3月に「Science」誌に発表された別の研究では、古典期マヤ文明が紀元800年頃に衰退した主な原因は干ばつであったと指摘されている。
「今回の研究結果を元に、今後いくつもの新たな研究が行われるものと確信している」と、スペイン、ジローナ大学のネウス・イサーン(Neus Isern)氏は言う。
イサーン氏は人口モデリングの専門家である。「背景に自然災害がなかったというのなら、新石器時代の経済はなぜ破綻したのか。新石器時代の経済は、私たちが考えるほど持続的なものではなかったのだろうか?」
ダウニー氏が推測しているのは、初期農耕民族は森林の乱伐や農地の継続使用によって急速な土壌の荒廃を招き、その結果、疾病が流行しやすくなって、人口減少の引き金を引いたというシナリオだ。
もう1つの可能性として、アイルランドにあるユニバーシティ・カレッジ・コークの考古学者ロン・ピンハシ(Ron Pinhasi)氏が提唱するのは、人口の大幅な増減の主な原因を移住であるとする考え方だ。
「2度目の人口増加は、乳製品のような二次生産物が作られはじめるという革新の結果であった可能性がある。酪農じたいは以前から始まっていたが、各地に広がるのは後になってからだ」とピンハシ氏は言う。
一般的にヒトの成人は牛乳などに含まれる乳糖を腸で分解できないが、ヨーロッパでは遺伝子変異のおかげで、90%近い成人が牛乳を飲める。
2009年の研究によると、この遺伝子変異はわずか7500年ほど前に起こった可能性が高いという。
「この研究結果には、現代一般的な考え方に対する、非常に重要なメッセージが含まれていると思わずにいられない。
現代では、新たなテクノロジーこそが常に時代を制するという盲信があちこちに見られるが、それは新石器時代には当てはまらないのだから」とダウニー氏は言う。
ヨーロッパの初期農耕時代に関する今回の研究は、「Nature Communications」誌に10月1日に掲載された。
552 :世界@名無史さん:2013/11/17(日) 14:56:09.55 0
この酪農をもたらしたのが遺伝子で言うとR1aなのだろう。
言語的には印欧語族。
そして人種的には金髪碧眼。
古代からのクロマニヨン人の子孫や南方からの農耕民族に4〜5000年前に取って代わった連中だ。
欧州の古代史もようやく着地点が見えてきた。
言語的には印欧語族。
そして人種的には金髪碧眼。
古代からのクロマニヨン人の子孫や南方からの農耕民族に4〜5000年前に取って代わった連中だ。
欧州の古代史もようやく着地点が見えてきた。
欧州の古代史の着地点が幻となって現れたの間違いでは?
【人類】「民族として歴史が浅いかもしれない?」…ヨーロッパ人の遺伝学的な起源が判明
http://anago.2ch.net/test/read.cgi/scienceplus/1367103944/1
http://anago.2ch.net/test/read.cgi/scienceplus/1367103944/1
>>554
1 名前:ケンシロウとユリア百式φ ★[] 投稿日:2013/04/28(日) 08:05:44.29 ID:???
現代ヨーロッパの遺伝学的な歴史が初めて詳述された。
最新の研究によれば、民族としてのヨーロッパ人は予想以上に歴史が浅いかもしれないという。
古代の骨から採取したDNAを調べた結果、現代ヨーロッパ人の遺伝子構造は6500年前、
新石器時代の中期にあたる紀元前4500年頃に定着したことがわかった。
約7500年前にやって来た最初の農耕民や、さらに古い時代の狩猟採集民がもたらしたものではなかった。
研究を実施したアデレード大学オーストラリア古代DNAセンター
(Australian Centre for Ancient DNA)の所長アラン・クーパー(Alan Cooper)氏は、
「原因は不明だが当時、住んでいた人たちの遺伝的な特性が消失したらしい」と説明する。
この意外な調査結果は新しい研究の一部で、現代ヨーロッパの遺伝的な歴史を初めて詳述している。
共同研究者で、ナショナル ジオグラフィック協会のジェノグラフィック・プロジェクトを率いる
遺伝学者スペンサー・ウェルズ(Spencer Wells)氏は、
「比較的新しい時代の民族移動が、中欧の人々に遺伝学的に重大な影響を及ぼしたようだ」と話す。
(>>2以降に続きます)[1/3]
ソース:ナショナルジオグラフィック(April 25, 2013)
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=20130425003
画像:ドイツ、ブライスガウ出身者で編成された部隊。1920年代に撮影
http://www.nationalgeographic.co.jp/news/news_images/66603_0_600x450.jpg
関連リンク:naturecommunicationsに掲載された論文要旨
「Neolithic mitochondrial haplogroup H genomes and the genetic origins of Europeans」(英文)
http://www.nature.com/ncomms/journal/v4/n4/full/ncomms2656.html
1 名前:ケンシロウとユリア百式φ ★[] 投稿日:2013/04/28(日) 08:05:44.29 ID:???
現代ヨーロッパの遺伝学的な歴史が初めて詳述された。
最新の研究によれば、民族としてのヨーロッパ人は予想以上に歴史が浅いかもしれないという。
古代の骨から採取したDNAを調べた結果、現代ヨーロッパ人の遺伝子構造は6500年前、
新石器時代の中期にあたる紀元前4500年頃に定着したことがわかった。
約7500年前にやって来た最初の農耕民や、さらに古い時代の狩猟採集民がもたらしたものではなかった。
研究を実施したアデレード大学オーストラリア古代DNAセンター
(Australian Centre for Ancient DNA)の所長アラン・クーパー(Alan Cooper)氏は、
「原因は不明だが当時、住んでいた人たちの遺伝的な特性が消失したらしい」と説明する。
この意外な調査結果は新しい研究の一部で、現代ヨーロッパの遺伝的な歴史を初めて詳述している。
共同研究者で、ナショナル ジオグラフィック協会のジェノグラフィック・プロジェクトを率いる
遺伝学者スペンサー・ウェルズ(Spencer Wells)氏は、
「比較的新しい時代の民族移動が、中欧の人々に遺伝学的に重大な影響を及ぼしたようだ」と話す。
(>>2以降に続きます)[1/3]
ソース:ナショナルジオグラフィック(April 25, 2013)
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=20130425003
画像:ドイツ、ブライスガウ出身者で編成された部隊。1920年代に撮影
http://www.nationalgeographic.co.jp/news/news_images/66603_0_600x450.jpg
{kind=link}
関連リンク:naturecommunicationsに掲載された論文要旨
「Neolithic mitochondrial haplogroup H genomes and the genetic origins of Europeans」(英文)
http://www.nature.com/ncomms/journal/v4/n4/full/ncomms2656.html
>>554
◆遺伝的な特性
クーパー氏の研究チームは、ドイツ中央部で発掘された遺骨39体の歯や骨を研究対象に選んだ。
時代はおよそ7500年前から2500年前と推定されている。遺骨から、母親のみから受け継がれる遺伝情報、ミトコンドリアDNAを抽出。
次に、ミトコンドリアDNAの変異がよく似た1つのグループに焦点を当てた。
現代ヨーロッパ人の最大45%が受け継ぐ遺伝的集団、ハプログループHだ。
生物の遺伝的な構成(DNA配列)をハプロタイプと呼び、グループ分けした集団をハプログループと称す。
親から子に引き継がれるハプロタイプを調査すれば、人類集団の比較調査が可能になる。
ハプログループHの変異は、何千年も前からヨーロッパ人の遺伝子構造に存在した可能性が過去の研究によって示されている。
この集団がヨーロッパで優勢になった経緯はわかっていない。
最後の氷河期が終了した約1万2000年前から人口が急増し、ヨーロッパ大陸に広がったのではないかという説もある。
しかし、クーパー氏らによる最新のデータは、現代ヨーロッパの遺伝的な基盤について異なる姿を描き出している。
1度や2度の移動ではなく、複数の集団が異なる方向からさまざまなタイミングで波のように押し寄せ、
ヨーロッパの人種構成は数度にわたって一変したというのだ。
最初の現生人類は3万5000〜4万年前、アフリカからヨーロッパにやって来た。
3万年前ごろまでにはヨーロッパ全域に広がり、一方、近縁種のネアンデルタール人は消失。
これら狩猟採集民のDNAが、ハプログループHだった可能性は低い。
約7500年前の新石器時代前期、今度は中東からヨーロッパに次の波が押し寄せた。
作物の栽培方法とともに、ハプログループHの変異型が持ち込まれた。
考古学者たちは、中欧にやって来た最初の農耕民が形成した文化を線形陶器文化(LBK)と呼んでいる。
線形の装飾を施した陶器が多く見つかっているためだ。
遺伝的な証拠によって、LBKの農耕民と彼ら固有のハプログループHの到来が、ハプログループUの激減と
一致することが示されている。当時、ヨーロッパの狩猟採集民の間では、ハプログループUが優勢だった。
◆遺伝的な特性
クーパー氏の研究チームは、ドイツ中央部で発掘された遺骨39体の歯や骨を研究対象に選んだ。
時代はおよそ7500年前から2500年前と推定されている。遺骨から、母親のみから受け継がれる遺伝情報、ミトコンドリアDNAを抽出。
次に、ミトコンドリアDNAの変異がよく似た1つのグループに焦点を当てた。
現代ヨーロッパ人の最大45%が受け継ぐ遺伝的集団、ハプログループHだ。
生物の遺伝的な構成(DNA配列)をハプロタイプと呼び、グループ分けした集団をハプログループと称す。
親から子に引き継がれるハプロタイプを調査すれば、人類集団の比較調査が可能になる。
ハプログループHの変異は、何千年も前からヨーロッパ人の遺伝子構造に存在した可能性が過去の研究によって示されている。
この集団がヨーロッパで優勢になった経緯はわかっていない。
最後の氷河期が終了した約1万2000年前から人口が急増し、ヨーロッパ大陸に広がったのではないかという説もある。
しかし、クーパー氏らによる最新のデータは、現代ヨーロッパの遺伝的な基盤について異なる姿を描き出している。
1度や2度の移動ではなく、複数の集団が異なる方向からさまざまなタイミングで波のように押し寄せ、
ヨーロッパの人種構成は数度にわたって一変したというのだ。
最初の現生人類は3万5000〜4万年前、アフリカからヨーロッパにやって来た。
3万年前ごろまでにはヨーロッパ全域に広がり、一方、近縁種のネアンデルタール人は消失。
これら狩猟採集民のDNAが、ハプログループHだった可能性は低い。
約7500年前の新石器時代前期、今度は中東からヨーロッパに次の波が押し寄せた。
作物の栽培方法とともに、ハプログループHの変異型が持ち込まれた。
考古学者たちは、中欧にやって来た最初の農耕民が形成した文化を線形陶器文化(LBK)と呼んでいる。
線形の装飾を施した陶器が多く見つかっているためだ。
遺伝的な証拠によって、LBKの農耕民と彼ら固有のハプログループHの到来が、ハプログループUの激減と
一致することが示されている。当時、ヨーロッパの狩猟採集民の間では、ハプログループUが優勢だった。
>>554
◆農耕民の到来
今回の発見は、専門家の間で長年続く論争を解決に導くと、ウェルズ氏は話す。
同氏はナショナル ジオグラフィック協会付き探検家でもある。
「ヨーロッパの考古学的な記録には遺伝子変化を伴う。つまり、人とそのDNAの移動に伴って、文化の変化が起きたのだ」。
LBKグループとその子孫は繁栄を極め、瞬く間にヨーロッパ全域に広がった。
「ヨーロッパを席巻した最初の文化と言ってもよいだろう」とクーパー氏は話す。
この大躍進を考えると、彼らが多くの現代ヨーロッパ人の重要な遺伝的祖先と考えるのが自然だ。
ところが研究チームの遺伝子解析結果は、常識を覆すような事実を示している。
約6500年前の新石器時代中期、LBKの文化が衰退。そして突然、このグループ固有のハプログループH変異型が
希少な存在となり、同グループの異なる変異型を持つ人々に取って代わられてしまうのだ。
今回の研究結果は、オンライン科学誌「Nature Communications」に4月23日付けで発表されている。
◆農耕民の到来
今回の発見は、専門家の間で長年続く論争を解決に導くと、ウェルズ氏は話す。
同氏はナショナル ジオグラフィック協会付き探検家でもある。
「ヨーロッパの考古学的な記録には遺伝子変化を伴う。つまり、人とそのDNAの移動に伴って、文化の変化が起きたのだ」。
LBKグループとその子孫は繁栄を極め、瞬く間にヨーロッパ全域に広がった。
「ヨーロッパを席巻した最初の文化と言ってもよいだろう」とクーパー氏は話す。
この大躍進を考えると、彼らが多くの現代ヨーロッパ人の重要な遺伝的祖先と考えるのが自然だ。
ところが研究チームの遺伝子解析結果は、常識を覆すような事実を示している。
約6500年前の新石器時代中期、LBKの文化が衰退。そして突然、このグループ固有のハプログループH変異型が
希少な存在となり、同グループの異なる変異型を持つ人々に取って代わられてしまうのだ。
今回の研究結果は、オンライン科学誌「Nature Communications」に4月23日付けで発表されている。
http://yuihaga.blog.fc2.com/blog-entry-238.html
オックスフォード大学(英国)の考古学者、トム・ハイアム(Tom Higham)が率いるチームは、
骨の中のコラーゲンタンパクからコンタミネーションを除去する手法を使ってヨーロッパ中の何十か所もの人類とネアンデルタール人の遺跡を再年代測定している。
このプロジェクトは人類が40,000年以上前にブリテンに達した――以前の考えより約10,000年早い――ことを発見し、
抽象的思考の能力を示す貝殻ビーズなどの物をネアンデルタール人が作った証拠に異議を唱えている。
オックスフォード大学(英国)の考古学者、トム・ハイアム(Tom Higham)が率いるチームは、
骨の中のコラーゲンタンパクからコンタミネーションを除去する手法を使ってヨーロッパ中の何十か所もの人類とネアンデルタール人の遺跡を再年代測定している。
このプロジェクトは人類が40,000年以上前にブリテンに達した――以前の考えより約10,000年早い――ことを発見し、
抽象的思考の能力を示す貝殻ビーズなどの物をネアンデルタール人が作った証拠に異議を唱えている。
Europeans and South Asians share by descent SLC24A5 light skin allele
http://dienekes.blogspot.jp/2013/11/europeans-and-south-asians-share-by.html
The age estimate for this allele is quite old but with a huge 95% confidence interval.
Hopefully ancient DNA can illuminate the trajectory of the allele's frequency through time and space.
http://2.bp.blogspot.com/-rhqReVApmRk/Un1LdnrSSyI/AAAAAAAAJY8/cmRVUA1On9k/s1600/journal.pgen.1003912.g002.png
Sequencing 11.74 kb of SLC24A5 in 95 individuals worldwide reveals that the rs1426654-A alleles in South Asian and West Eurasian populations
are monophyletic and occur on the background of a common haplotype that is characterized by low genetic diversity.
We date the coalescence of the light skin associated allele at 22?28 KYA.
Both our sequence and genome-wide genotype data confirm that this gene has been a target for positive selection among Europeans.
However, the latter also shows additional evidence of selection in populations of the Middle East, Central Asia, Pakistan and North India but not in South India.
http://dienekes.blogspot.jp/2013/11/europeans-and-south-asians-share-by.html
The age estimate for this allele is quite old but with a huge 95% confidence interval.
Hopefully ancient DNA can illuminate the trajectory of the allele's frequency through time and space.
http://2.bp.blogspot.com/-rhqReVApmRk/Un1LdnrSSyI/AAAAAAAAJY8/cmRVUA1On9k/s1600/journal.pgen.1003912.g002.png
{kind=link}
Sequencing 11.74 kb of SLC24A5 in 95 individuals worldwide reveals that the rs1426654-A alleles in South Asian and West Eurasian populations
are monophyletic and occur on the background of a common haplotype that is characterized by low genetic diversity.
We date the coalescence of the light skin associated allele at 22?28 KYA.
Both our sequence and genome-wide genotype data confirm that this gene has been a target for positive selection among Europeans.
However, the latter also shows additional evidence of selection in populations of the Middle East, Central Asia, Pakistan and North India but not in South India.
560 :世界@名無史さん:2013/12/07(土) 10:27:38.40 0
欧州の歴史もようやく遺伝子の歴史的変化を反映させるものになるのかな。
クロマニヨン人の子孫なんかごく一部だよ。
過半は東方からの移住者。
クロマニヨン人の子孫なんかごく一部だよ。
過半は東方からの移住者。
561 :世界@名無史さん:2013/12/07(土) 10:49:32.07 0
黄色人種でも、在特会の桜井誠のような人間は容姿端麗だな。
欧米人以上だ。
欧米人以上だ。
563 :世界@名無史さん:2013/12/07(土) 14:38:31.98 0
桜井誠ってぐぐってみたけど、
画像見たらキモヲタじゃねーか!!!
画像見たらキモヲタじゃねーか!!!
564 :世界@名無史さん:2013/12/08(日) 08:58:49.76 0
>>562
Y遺伝子の話。
クロマニヨン⇒I
農耕をもたらした民族⇒EorJ
この4000〜5000年前に東方から侵入した印欧語族⇒R1aorR1b
こういう関係なんだが、ようやく学会でも公認され始めた。
Y遺伝子の話。
クロマニヨン⇒I
農耕をもたらした民族⇒EorJ
この4000〜5000年前に東方から侵入した印欧語族⇒R1aorR1b
こういう関係なんだが、ようやく学会でも公認され始めた。
えーっと、そんなもんすかね
http://en.wikipedia.org/wiki/Cro-Magnon
Current scientific literature prefers the term European Early Modern Humans (EEMH),
to the term 'Cro-Magnon,' which has no formal taxonomic status, as it refers neither to a species or subspecies nor to an archaeological phase or culture.[1]
The earliest known remains of Cro-Magnon-like humans are radiocarbon dated to 43,000 years before present.[2]
http://en.wikipedia.org/wiki/File:Cro-Magnon_range_37,500_ybp.svg
http://en.wikipedia.org/wiki/File:Cro-Magnon_range_35,000_ybp.svg
http://en.wikipedia.org/wiki/File:Cro-Magnon_range_32,500_ybp.svg
http://en.wikipedia.org/wiki/File:Cro-Magnon_range_30,000_ybp.svg
http://en.wikipedia.org/wiki/Cro-Magnon
Current scientific literature prefers the term European Early Modern Humans (EEMH),
to the term 'Cro-Magnon,' which has no formal taxonomic status, as it refers neither to a species or subspecies nor to an archaeological phase or culture.[1]
The earliest known remains of Cro-Magnon-like humans are radiocarbon dated to 43,000 years before present.[2]
http://en.wikipedia.org/wiki/File:Cro-Magnon_range_37,500_ybp.svg
{kind=link}
http://en.wikipedia.org/wiki/File:Cro-Magnon_range_35,000_ybp.svg
{kind=link}
http://en.wikipedia.org/wiki/File:Cro-Magnon_range_32,500_ybp.svg
{kind=link}
http://en.wikipedia.org/wiki/File:Cro-Magnon_range_30,000_ybp.svg
{kind=link}
Y-DNA Haplogroup I Naming Nomenclature
http://www.genebase.com/learning/article/60
Learn about Y-chromosome Haplogroup I
http://www.genebase.com/learning/article/12
Learn about Y-DNA Haplogroup R
http://www.genebase.com/learning/article/11
残念ながらUの方はみつかんないね
最重要なのに
Learn about mtDNA Haplogroup H
http://www.genebase.com/learning/article/20
http://www.genebase.com/learning/article/60
Learn about Y-chromosome Haplogroup I
http://www.genebase.com/learning/article/12
Learn about Y-DNA Haplogroup R
http://www.genebase.com/learning/article/11
残念ながらUの方はみつかんないね
最重要なのに
Learn about mtDNA Haplogroup H
http://www.genebase.com/learning/article/20
>>564
詳しそうだから聞きます
>>558のブリテンに到達した4万年前の現世人類(おそらくクロマニヨン)って
どの個体の話なんでしょうか?
見つけれた今のところ一番古い記録は>>565にある33,000年前の男性っぽい
http://en.wikipedia.org/wiki/Red_Lady_of_Paviland
詳しそうだから聞きます
>>558のブリテンに到達した4万年前の現世人類(おそらくクロマニヨン)って
どの個体の話なんでしょうか?
見つけれた今のところ一番古い記録は>>565にある33,000年前の男性っぽい
http://en.wikipedia.org/wiki/Red_Lady_of_Paviland
>>568
すまんコれだ
http://en.wikipedia.org/wiki/Kents_Cavern
フランスの地図に見えたわ
ハプログループの表示は無さそうだね
>>568のはmtDNAのHみたいだけど
すまんコれだ
http://en.wikipedia.org/wiki/Kents_Cavern
フランスの地図に見えたわ
ハプログループの表示は無さそうだね
>>568のはmtDNAのHみたいだけど
>>566年代
Haplogroup R dates to 30-35kya in the Paleolithic Era and Pleistocene Epoch.
Glacial advances forced retreats of humans, including populations with Haplogroup R into refugia ?? one in Iberia (the R1b subclade) and one in Ukraine (forerunner R1a subclade).
The expansion of both populations occurred after the last glacial maximum (LGM) ~18kya.
R1b (~30kya) was a branch founded earlier than R1a (15-10kya).
R1a has been associated with the earliest Indo-European language populations and an early Asian culture, the Kurgan culture.
Haplogroup R1a has also spread to the east and reached Southeast Asia and Australia, albeit at modest levels (??10%).
Haplogroup R dates to 30-35kya in the Paleolithic Era and Pleistocene Epoch.
Glacial advances forced retreats of humans, including populations with Haplogroup R into refugia ?? one in Iberia (the R1b subclade) and one in Ukraine (forerunner R1a subclade).
The expansion of both populations occurred after the last glacial maximum (LGM) ~18kya.
R1b (~30kya) was a branch founded earlier than R1a (15-10kya).
R1a has been associated with the earliest Indo-European language populations and an early Asian culture, the Kurgan culture.
Haplogroup R1a has also spread to the east and reached Southeast Asia and Australia, albeit at modest levels (??10%).
571 :世界@名無史さん:2013/12/11(水) 19:59:42.70 0
>>566>>571
とりあえずRのクレードは>>559には関係がありそうに見えて色々と合わないな
そんでもってR1aって話だけどね
だが、まぁこっちはこのスレとは別の話だな
インドで生じた人種的混合の遺伝的証拠の発見、ハーバード大
http://jp.sciencenewsline.com/articles/2013080816380013.html
とりあえずRのクレードは>>559には関係がありそうに見えて色々と合わないな
そんでもってR1aって話だけどね
だが、まぁこっちはこのスレとは別の話だな
インドで生じた人種的混合の遺伝的証拠の発見、ハーバード大
http://jp.sciencenewsline.com/articles/2013080816380013.html
575 :世界@名無史さん:2013/12/12(木) 18:24:49.27 0
ミトコンドリアDNAのHはクロマニヨン人からも検出されてるけど、今存在するのとは系統が違うってこと?
577 :世界@名無史さん:2013/12/12(木) 21:42:29.09 0
去年あたりから、
古代欧州人は、
採集狩猟民⇒農耕民⇒酪農民
こういう主張が強くなっているね。
その根拠に遺伝子の変化が利用されている。
古代欧州人は、
採集狩猟民⇒農耕民⇒酪農民
こういう主張が強くなっているね。
その根拠に遺伝子の変化が利用されている。
578 :世界@名無史さん:2013/12/12(木) 21:45:07.88 0
神が造ったのが白人種で、
サルがアジア人と黒人。
これですべての説明がつくよ。
だから白人が「黒人とチンチャンチョンに
人権があるのなら、チンパンジーにだって人権を」というのさ。
サルがアジア人と黒人。
これですべての説明がつくよ。
だから白人が「黒人とチンチャンチョンに
人権があるのなら、チンパンジーにだって人権を」というのさ。
>>566そんで
Y-DNA Haplogroup I
とりあえ図
http://64.40.115.138/file/lu/6/52235/NTIyMzV9K3szNTcxNjU=.jpg?download=1
http://64.40.115.138/file/lu/6/52235/NTIyMzV9K3szNTcxNjY=.jpg?download=1
Figure 5. Proposed scenario of the movements of human populations bearing haplogroup I.
The figure shows likely origin of haplogroup I from the Middle East via the Levantine corridor and its residence in two pockets (gray ovals)
during the last glacial maximum in Europe. Subclades of haplogroup I are color coded and corresponding SNPs listed in the legend.
The routes and locations are based on evidence from several studies of Y-DNA I haplogroups and haplotypes.
この辺りまではなんとなく知ってるが、
サルディーニャが特別なのは意識してなかったね。
どおりで参照されるはずだわ
I2a2(M26)
http://64.40.115.138/file/lu/6/52235/NTIyMzV9K3szNTcxNjk=.jpg?download=1
Y-DNA Haplogroup I
とりあえ図
http://64.40.115.138/file/lu/6/52235/NTIyMzV9K3szNTcxNjU=.jpg?download=1
{kind=link}
http://64.40.115.138/file/lu/6/52235/NTIyMzV9K3szNTcxNjY=.jpg?download=1
{kind=link}
Figure 5. Proposed scenario of the movements of human populations bearing haplogroup I.
The figure shows likely origin of haplogroup I from the Middle East via the Levantine corridor and its residence in two pockets (gray ovals)
during the last glacial maximum in Europe. Subclades of haplogroup I are color coded and corresponding SNPs listed in the legend.
The routes and locations are based on evidence from several studies of Y-DNA I haplogroups and haplotypes.
この辺りまではなんとなく知ってるが、
サルディーニャが特別なのは意識してなかったね。
どおりで参照されるはずだわ
I2a2(M26)
http://64.40.115.138/file/lu/6/52235/NTIyMzV9K3szNTcxNjk=.jpg?download=1
{kind=link}
580 :世界@名無史さん:2013/12/12(木) 22:05:36.96 0
しかも、イタリア各州のなかでは黒目黒髪比率が最も高い。
やはりクロマニヨン人は黒目黒髪浅黒だったのだろう。
やはりクロマニヨン人は黒目黒髪浅黒だったのだろう。
>>579
I1. M253
The I1 subclade, defined by SNP M253 is prominent in Scandinavia and other Nordic countries, which includes Sweden (40-50%), Norway (40%), Finland (20%) and Denmark (40%).
It is the most prevalent I subhaplogroup in Sweden and TMRCA estimates are 6,000-8,000 years ago.
This is after the time when the glaciers are believed to have retreated from Fennoscandia.
The Saami population around Lappland in the northern regions of Fennoscandia also has high levels of this subclade.
I1 levels diminish moving in the eastern direction from Finland toward Russia and its frequency is low in other Eastern European regions (5-1%).
This and additional information from mtDNA haplogroups has provided evidence that the Saami came from a Southern European source rather than from Siberia and the East.
I1. M253
The I1 subclade, defined by SNP M253 is prominent in Scandinavia and other Nordic countries, which includes Sweden (40-50%), Norway (40%), Finland (20%) and Denmark (40%).
It is the most prevalent I subhaplogroup in Sweden and TMRCA estimates are 6,000-8,000 years ago.
This is after the time when the glaciers are believed to have retreated from Fennoscandia.
The Saami population around Lappland in the northern regions of Fennoscandia also has high levels of this subclade.
I1 levels diminish moving in the eastern direction from Finland toward Russia and its frequency is low in other Eastern European regions (5-1%).
This and additional information from mtDNA haplogroups has provided evidence that the Saami came from a Southern European source rather than from Siberia and the East.
>>581
The presence of haplogroup I1 in Scandinavia is linked with Viking (Norwegian or Danish) excursions in the North Atlantic
and the dissemination of haplogroup I1 in England, Ireland, Scotland and Iceland.
It is also found at high levels in Germany (~25%) and the Netherlands (~17%).
Subclade I1 is associated with the Anglo-Saxons who were believed to be descendents of Celtic and Germanic groups emanating
from the Low Countries and Frisia (areas near the Netherlands, Belgium, Luxembourg, Germany and Denmark).
Estimates of TMRCA for the Anglo-Saxon I1 subclade range from 1,500 to 4,000 years ago.
There is also speculation that the spread of Anglo-Saxons (and subclade I1) to England took place through a land bridge,
known as Doggerland that connected continental Europe to England around 10kya.
The presence of haplogroup I1 in Scandinavia is linked with Viking (Norwegian or Danish) excursions in the North Atlantic
and the dissemination of haplogroup I1 in England, Ireland, Scotland and Iceland.
It is also found at high levels in Germany (~25%) and the Netherlands (~17%).
Subclade I1 is associated with the Anglo-Saxons who were believed to be descendents of Celtic and Germanic groups emanating
from the Low Countries and Frisia (areas near the Netherlands, Belgium, Luxembourg, Germany and Denmark).
Estimates of TMRCA for the Anglo-Saxon I1 subclade range from 1,500 to 4,000 years ago.
There is also speculation that the spread of Anglo-Saxons (and subclade I1) to England took place through a land bridge,
known as Doggerland that connected continental Europe to England around 10kya.
>>579
I1a. M21
The I1a subclade can be described by the presence of SNP M21.
Currently little information exists concerning the frequency and distribution of this subclade.
I1b. M227
The M227 SNP identifies subclade I1b and appears to be a relatively new SNP mutation.
It is present at modest levels (0.5-2.0%) in Eastern Europe and the Balkans and is likely to be a minor subclade.
I1b1. M72
The M72 SNP identifies subclade I1b1.
Currently little information exists concerning the frequency and distribution of this subclade.
この辺りは適当だなww
Check this site regularly for updates on this subclade as new information will be posted as studies become available
http://www.eupedia.com/images/content/timeline_comparison.gif
http://www.eupedia.com/europe/Haplogroup_I1_Y-DNA.shtml
http://www.eupedia.com/images/content/I1-tree.gif
I1a. M21
The I1a subclade can be described by the presence of SNP M21.
Currently little information exists concerning the frequency and distribution of this subclade.
I1b. M227
The M227 SNP identifies subclade I1b and appears to be a relatively new SNP mutation.
It is present at modest levels (0.5-2.0%) in Eastern Europe and the Balkans and is likely to be a minor subclade.
I1b1. M72
The M72 SNP identifies subclade I1b1.
Currently little information exists concerning the frequency and distribution of this subclade.
この辺りは適当だなww
Check this site regularly for updates on this subclade as new information will be posted as studies become available
http://www.eupedia.com/images/content/timeline_comparison.gif
{kind=link}
http://www.eupedia.com/europe/Haplogroup_I1_Y-DNA.shtml
http://www.eupedia.com/images/content/I1-tree.gif
{kind=link}
>>579
I2
The I2 subclade is the group descended from the refugium centered in the Balkans and has maintained its predominance in this region,
which is marked by the Dinaric Alps running along the Adriatic Sea.
It is the most prevalent haplogroup in Bosnia, Herzegovinia, Croatia and Serbia.
It is also found abundantly on several islands in the Adriatic (Bra?, Hvar, Krk and Kor?ula) and Mediterranean (Sardinia) Seas.
Note that this is not a universal phenomenon since the corresponding I2 subclades are absent from Sicily and Corsica and below 5% on Crete.
It appears to be primarily associated with Slavic groups in Eastern Europe.
I2 subclade levels diminish to much lower levels in Western Europe and where they are associated with the Atlantic coastline.
I2
The I2 subclade is the group descended from the refugium centered in the Balkans and has maintained its predominance in this region,
which is marked by the Dinaric Alps running along the Adriatic Sea.
It is the most prevalent haplogroup in Bosnia, Herzegovinia, Croatia and Serbia.
It is also found abundantly on several islands in the Adriatic (Bra?, Hvar, Krk and Kor?ula) and Mediterranean (Sardinia) Seas.
Note that this is not a universal phenomenon since the corresponding I2 subclades are absent from Sicily and Corsica and below 5% on Crete.
It appears to be primarily associated with Slavic groups in Eastern Europe.
I2 subclade levels diminish to much lower levels in Western Europe and where they are associated with the Atlantic coastline.
>>584
I2a. P37.2
The I2a subclade can be described by the presence of SNP P37.2 and is the most frequent haplogroup in the Balkan region, with a frequency peaking in Herzegovinia (60-70%).
A high frequency is also found in Bosnia (40-50%) and Croatia (32%).
The Croatian islands frequencies (Hvar = 66%, Kor?ula 52%, Bra? =36% and Krk = 9.5%) show a decreasing trend in a northwestern direction.
A similar drop off is seen moving to the northeast through Russia.
In fact, the drop off in all directions creates a rather clear I2a peak in the Balkans that radiates outward over many Slavic populations to the east and south.
The I2a subclade divergence is estimated near 11kya
and it may have paralleled the spread of the R1a haplogroup in the Holocene period after the cold reversal and retreat of the glaciers from Europe.
I2a. P37.2
The I2a subclade can be described by the presence of SNP P37.2 and is the most frequent haplogroup in the Balkan region, with a frequency peaking in Herzegovinia (60-70%).
A high frequency is also found in Bosnia (40-50%) and Croatia (32%).
The Croatian islands frequencies (Hvar = 66%, Kor?ula 52%, Bra? =36% and Krk = 9.5%) show a decreasing trend in a northwestern direction.
A similar drop off is seen moving to the northeast through Russia.
In fact, the drop off in all directions creates a rather clear I2a peak in the Balkans that radiates outward over many Slavic populations to the east and south.
The I2a subclade divergence is estimated near 11kya
and it may have paralleled the spread of the R1a haplogroup in the Holocene period after the cold reversal and retreat of the glaciers from Europe.
>>584>>579
I2a2. M26
I2a2 is described by the presence of the M26 SNP and this interesting subclade of haplogroup I represents over 1/3 of the Y-chromosomes on the Mediterranean island of Sardinia.
Its particular pre-eminence here and low levels in only a few other Western European populations has led researchers to conclude that this subclade either originated on Sardinia or was part of its ancestral colonization.
Subsequently, in the absence of invasion and re-peopling, I2a2 became a founding and highly representative Y-chromosome on this island.
Another way to view this is that there may have been a ‘Sardinian haplogroup I Adam’ from which one out of three Sardinian males can trace their Y-chromosome.
Interestingly, the neighboring island of Corsica lacks this subclade altogether and appears to have an overall low frequency of haplogroup I.
This could have been due to a smaller initial population of I2a2 coupled with successful colonization by other populations over time and replacement of this unique I subclade.
An estimate for the I2a2 Sardinian subclade TMRCA is ~21kya and expansion of the population around 14kya.
I2a2. M26
I2a2 is described by the presence of the M26 SNP and this interesting subclade of haplogroup I represents over 1/3 of the Y-chromosomes on the Mediterranean island of Sardinia.
Its particular pre-eminence here and low levels in only a few other Western European populations has led researchers to conclude that this subclade either originated on Sardinia or was part of its ancestral colonization.
Subsequently, in the absence of invasion and re-peopling, I2a2 became a founding and highly representative Y-chromosome on this island.
Another way to view this is that there may have been a ‘Sardinian haplogroup I Adam’ from which one out of three Sardinian males can trace their Y-chromosome.
Interestingly, the neighboring island of Corsica lacks this subclade altogether and appears to have an overall low frequency of haplogroup I.
This could have been due to a smaller initial population of I2a2 coupled with successful colonization by other populations over time and replacement of this unique I subclade.
An estimate for the I2a2 Sardinian subclade TMRCA is ~21kya and expansion of the population around 14kya.
>>584
I2b1. M223
The M223 SNP is a marker for the I2b1 subclade, although it has been linked with the I1 subclade by geographical distribution.
This subclade has been detected as a modest fraction of Y-chromosome haplogroup I in Sweden (5% of 42% total I).
A similar trend is found throughout Europe where it contributes between 1 and 25% of the total pool of haplogroup I.
Its highest frequency is located around Denmark, Germany and the Netherlands.
It was previously assigned as subclade I1b2a (also known as I1c in older studies), but the present phylogenetic tree from ISOGG places this group in the I2b subclade.
I2b1. M223
The M223 SNP is a marker for the I2b1 subclade, although it has been linked with the I1 subclade by geographical distribution.
This subclade has been detected as a modest fraction of Y-chromosome haplogroup I in Sweden (5% of 42% total I).
A similar trend is found throughout Europe where it contributes between 1 and 25% of the total pool of haplogroup I.
Its highest frequency is located around Denmark, Germany and the Netherlands.
It was previously assigned as subclade I1b2a (also known as I1c in older studies), but the present phylogenetic tree from ISOGG places this group in the I2b subclade.
>>587
I2b1. M223
9000ybp
The M223 SNP is a marker for the I2b1 subclade, although it has been linked with the I1 subclade by geographical distribution.
This subclade has been detected as a modest fraction of Y-chromosome haplogroup I in Sweden (5% of 42% total I).
A similar trend is found throughout Europe where it contributes between 1 and 25% of the total pool of haplogroup I.
Its highest frequency is located around Denmark, Germany and the Netherlands.
It was previously assigned as subclade I1b2a (also known as I1c in older studies), but the present phylogenetic tree from ISOGG places this group in the I2b subclade.
I2b1a. M284
2000ybp
The M284 SNP is a marker for the I2b1a subclade and may have arisen in Britain or Scotland where it appears to be uniquely found.
The large study of ~1,000 Europeans by Rootsi did not identify anyone with this variation.
There is little additional information concerning the frequency and distribution of this subclade.
I2b1b. M379
The I2b1b subclade can be described by the presence of SNP M379.
Currently, little information exists concerning the frequency and distribution of this subclade.
後は省略したいが、、、
http://www.eupedia.com/images/content/timeline_comparison.gif
I2b1. M223
9000ybp
The M223 SNP is a marker for the I2b1 subclade, although it has been linked with the I1 subclade by geographical distribution.
This subclade has been detected as a modest fraction of Y-chromosome haplogroup I in Sweden (5% of 42% total I).
A similar trend is found throughout Europe where it contributes between 1 and 25% of the total pool of haplogroup I.
Its highest frequency is located around Denmark, Germany and the Netherlands.
It was previously assigned as subclade I1b2a (also known as I1c in older studies), but the present phylogenetic tree from ISOGG places this group in the I2b subclade.
I2b1a. M284
2000ybp
The M284 SNP is a marker for the I2b1a subclade and may have arisen in Britain or Scotland where it appears to be uniquely found.
The large study of ~1,000 Europeans by Rootsi did not identify anyone with this variation.
There is little additional information concerning the frequency and distribution of this subclade.
I2b1b. M379
The I2b1b subclade can be described by the presence of SNP M379.
Currently, little information exists concerning the frequency and distribution of this subclade.
後は省略したいが、、、
http://www.eupedia.com/images/content/timeline_comparison.gif
>>585
I2a1a. P41.2
8000ybp
This subclade can be identified by the presence of SNP P41.2.
A very minor frequency (1/523 males or <1.0%) of this subclade has been detected in Turkey (Anatolia).
There is little additional information concerning the frequency and distribution of this subclade.
I2a2a. M161
7000ybp
This subclade can be described by the presence of SNP M161.
Currently, little information exists concerning the frequency and distribution of this subclade.
I2a1a. P41.2
8000ybp
This subclade can be identified by the presence of SNP P41.2.
A very minor frequency (1/523 males or <1.0%) of this subclade has been detected in Turkey (Anatolia).
There is little additional information concerning the frequency and distribution of this subclade.
I2a2a. M161
7000ybp
This subclade can be described by the presence of SNP M161.
Currently, little information exists concerning the frequency and distribution of this subclade.
591 :世界@名無史さん:2013/12/12(木) 23:17:37.29 0
IもR1bもここ数千年の間にグループ分かれしているね。
例えばI1なんか誕生は10000年以上前でも、
現在のI1は7000年前前後のたった1人の男性が共通先祖というのが定説化している。
R1bも西欧のなかでもいくつかの大きなグループに分かれ、
ケルト人など古代の民族と何らかの相関関係を有している。
まだまだこれからだよ遺伝人類学というか遺伝歴史学は。
例えばI1なんか誕生は10000年以上前でも、
現在のI1は7000年前前後のたった1人の男性が共通先祖というのが定説化している。
R1bも西欧のなかでもいくつかの大きなグループに分かれ、
ケルト人など古代の民族と何らかの相関関係を有している。
まだまだこれからだよ遺伝人類学というか遺伝歴史学は。
595 :世界@名無史さん:2013/12/13(金) 06:32:53.86 0
ジェファーソンのY-DNAのTはいつごろ入ってきたんだろう。アフリカの角に多いタイプだそうだけど
>The affiliation of the Jefferson haplotype to T1a* and the absence of closely related haplotypes (zero to two step mutations away) in the network
supports the hypothesis that this haplotype belongs to an ancient rare European Y-chromosome lineage rather than to lineages that recently migrated to Europe from the Near East.
ジェファーソンのハプロタイプがT1a*に属し、ネットワーク内での近縁のハプロタイプ(0から2段階の変異)が存在しないことは、
このハプロタイプが最近中東からヨーロッパに移住してきた系統というよりも、古い稀なヨーロッパのY染色体の系統に属するという仮説を支持するものである
英語版Wikipedia Haplogroup T-M184 内 Notable haplogroup membersより
出典らしき
http://www.bioone.org/doi/abs/10.3378/027.083.0103
ここではログインしないとまとめしか見られないので、上記の文が見当たらないけど、新石器時代の農耕民としてやってきたということでいいのかな
少なくともジェファーソン一族のはもっと古い系統ということもあり得るのかな
>The affiliation of the Jefferson haplotype to T1a* and the absence of closely related haplotypes (zero to two step mutations away) in the network
supports the hypothesis that this haplotype belongs to an ancient rare European Y-chromosome lineage rather than to lineages that recently migrated to Europe from the Near East.
ジェファーソンのハプロタイプがT1a*に属し、ネットワーク内での近縁のハプロタイプ(0から2段階の変異)が存在しないことは、
このハプロタイプが最近中東からヨーロッパに移住してきた系統というよりも、古い稀なヨーロッパのY染色体の系統に属するという仮説を支持するものである
英語版Wikipedia Haplogroup T-M184 内 Notable haplogroup membersより
出典らしき
http://www.bioone.org/doi/abs/10.3378/027.083.0103
ここではログインしないとまとめしか見られないので、上記の文が見当たらないけど、新石器時代の農耕民としてやってきたということでいいのかな
少なくともジェファーソン一族のはもっと古い系統ということもあり得るのかな
596 :世界@名無史さん:2013/12/13(金) 17:25:05.81 0
中東→近東に訂正
597 :世界@名無史さん:2013/12/13(金) 17:36:43.46 0
白人が容姿端麗というより、容姿端麗な白人はさほど大きく見えず人種的に近く見える。
綺麗な子だけ並べると人類に人種などは存在せず、単に顔立ちとメラニン色素量の個人差があるだけに見える。
ところが白人と黄色人種をブサイク同士並べると、明らかに白人の方が全パーツが大きく、
毛深い場合も多く成長スピードや肥満になるのも速く見え、黄色人種と全く異なる人種に思える。
綺麗な子だけ並べると人類に人種などは存在せず、単に顔立ちとメラニン色素量の個人差があるだけに見える。
ところが白人と黄色人種をブサイク同士並べると、明らかに白人の方が全パーツが大きく、
毛深い場合も多く成長スピードや肥満になるのも速く見え、黄色人種と全く異なる人種に思える。
さて、ごく自然に東から来たと仮定してみる
>>574
Upper Palaeolithic Siberian genome reveals dual ancestry of Native Americans
http://www.nature.com/nature/journal/vaop/ncurrent/abs/nature12736.html
MA-1
・Here we sequence the draft genome of an approximately 24,000-year-old individual (MA-1), from Mal’ta in south-central Siberi
・The MA-1 mitochondrial genome belongs to haplogroup U, which has also been found at high frequency among Upper Palaeolithic and Mesolithic European hunter-gatherers
・Y chromosome of MA-1 is basal to modern-day western Eurasians and near the root of most Native American lineages
・we estimate that 14 to 38% of Native American ancestry may originate through gene flow from this ancient population
Afontova Gora-2
・Sequencing of another south-central Siberian, Afontova Gora-2 dating to approximately 17,000 years ago,
・ revealed similar autosomal genetic signatures as MA-1
>>574
Upper Palaeolithic Siberian genome reveals dual ancestry of Native Americans
http://www.nature.com/nature/journal/vaop/ncurrent/abs/nature12736.html
MA-1
・Here we sequence the draft genome of an approximately 24,000-year-old individual (MA-1), from Mal’ta in south-central Siberi
・The MA-1 mitochondrial genome belongs to haplogroup U, which has also been found at high frequency among Upper Palaeolithic and Mesolithic European hunter-gatherers
・Y chromosome of MA-1 is basal to modern-day western Eurasians and near the root of most Native American lineages
・we estimate that 14 to 38% of Native American ancestry may originate through gene flow from this ancient population
Afontova Gora-2
・Sequencing of another south-central Siberian, Afontova Gora-2 dating to approximately 17,000 years ago,
・ revealed similar autosomal genetic signatures as MA-1
>>599
とりあえずmtDNAのサブクレードルは必須
Y-DNAの方もおまけとして調べてみることにしたい
http://dienekes.blogspot.jp/2013/11/ancient-dna-from-upper-paleolithic-lake.html
http://2.bp.blogspot.com/-Va8sH8ho66k/Uo0OKBpX4fI/AAAAAAAAJZo/RLd9i4JYpv4/s1600/pca.png
とりあえずmtDNAのサブクレードルは必須
Y-DNAの方もおまけとして調べてみることにしたい
http://dienekes.blogspot.jp/2013/11/ancient-dna-from-upper-paleolithic-lake.html
http://2.bp.blogspot.com/-Va8sH8ho66k/Uo0OKBpX4fI/AAAAAAAAJZo/RLd9i4JYpv4/s1600/pca.png
{kind=link}
>>600
さきにおまけから
http://sicambre.at.webry.info/201311/article_26.html
http://web.joumon.jp.net/blog/2008/11/000656.html
さきにおまけから
http://sicambre.at.webry.info/201311/article_26.html
http://web.joumon.jp.net/blog/2008/11/000656.html
>>599
>>600
データを見る限りMA-1
mtDNA U*
Y-DNA R*
に見えるな。
R1*と予想していただけにちょっと予想外
Today, hg U is a pan western Eurasian haplogroup, with distribution across Europe, the
Middle East, South and Central Asia, western Siberia and North Africa (Figure SI 4b).
The overall frequency of hg U is low or absent in extant central and south
Siberian populations, i.e. the region close to where MA-1 originated, as well as in
East Asia (Figure SI 4b).
>>600
データを見る限りMA-1
mtDNA U*
Y-DNA R*
に見えるな。
R1*と予想していただけにちょっと予想外
Today, hg U is a pan western Eurasian haplogroup, with distribution across Europe, the
Middle East, South and Central Asia, western Siberia and North Africa (Figure SI 4b).
The overall frequency of hg U is low or absent in extant central and south
Siberian populations, i.e. the region close to where MA-1 originated, as well as in
East Asia (Figure SI 4b).
603 :世界@名無史さん:2013/12/14(土) 18:14:15.17 0
RはQと枝分かれしたのが西シベリア南部から西トルキスタン西部にかけて。
RはそこからR1とR2に分化している。
だからR2はインド北西部中心の分布になっている。
R1の一部はQのように米大陸に渡っている(北米東部に分布)。
そこからR1aとR1bに分化している。これが1万5千年より少し前ぐらい。
そこからR1a・R1bともにあちこちへ移動している。
R1bの一部は分化初期に何故か中近東を突っ切ってアフリカのカメルーンへ到達している。
R1bの主力が欧州へ本格侵入したのは5千年ぐらい前のことだよ。
RはそこからR1とR2に分化している。
だからR2はインド北西部中心の分布になっている。
R1の一部はQのように米大陸に渡っている(北米東部に分布)。
そこからR1aとR1bに分化している。これが1万5千年より少し前ぐらい。
そこからR1a・R1bともにあちこちへ移動している。
R1bの一部は分化初期に何故か中近東を突っ切ってアフリカのカメルーンへ到達している。
R1bの主力が欧州へ本格侵入したのは5千年ぐらい前のことだよ。
604 :世界@名無史さん:2013/12/14(土) 22:28:12.46 0
>R1bの一部は分化初期に何故か中近東を突っ切ってアフリカのカメルーンへ到達している。
ラムセス2世のY−DNAのタイプがE系統(なぜか中東系と言うよりもサブサハラのタイプ)で、一方ツタンカーメンはR1bだから
この二人の治世の間のどこかの時期と言うことになるな。カメルーンのR1bを持つグループには、何か出自に関する伝説とかあるんだろうか
ラムセス2世のY−DNAのタイプがE系統(なぜか中東系と言うよりもサブサハラのタイプ)で、一方ツタンカーメンはR1bだから
この二人の治世の間のどこかの時期と言うことになるな。カメルーンのR1bを持つグループには、何か出自に関する伝説とかあるんだろうか
605 :世界@名無史さん:2013/12/15(日) 09:46:46.25 0
>>604
良い質問だね。
中東・北アフリカの古代史解明の手掛かりのひとつになる。
仮説1:R1bが中東から移動中に随所に子孫を残したとして、その系統。
仮説2:ツタンカーメンは養子。出身一族がヒッタイトなどの印欧語族の系統。
個人的には仮説2じゃないかと思ってるけど根拠はない。
ただ、アンケセナーメンが夫の死後、ヒッタイト国王に自分の新夫をくれと手紙を出している。
だから、エジプトの上層階級は印欧語族と何らかの血縁関係があったのかもしれない。
良い質問だね。
中東・北アフリカの古代史解明の手掛かりのひとつになる。
仮説1:R1bが中東から移動中に随所に子孫を残したとして、その系統。
仮説2:ツタンカーメンは養子。出身一族がヒッタイトなどの印欧語族の系統。
個人的には仮説2じゃないかと思ってるけど根拠はない。
ただ、アンケセナーメンが夫の死後、ヒッタイト国王に自分の新夫をくれと手紙を出している。
だから、エジプトの上層階級は印欧語族と何らかの血縁関係があったのかもしれない。
いや、単にアッシリアがミタンニを陥落させたので同盟を強化したかったんだろ
>>559>>574>>599>>602
検証したところ特に問題はないだろうな
とりあえずバクトリアの辺りが
アレキサンダー以後か以前かは調べてみたい気はする
>>573調べたらある程度わかるんだろうね
検証したところ特に問題はないだろうな
とりあえずバクトリアの辺りが
アレキサンダー以後か以前かは調べてみたい気はする
>>573調べたらある程度わかるんだろうね
608 :世界@名無史さん:2013/12/16(月) 20:05:58.70 0
>>559
これは最新の論文かな。
よく読んでみよう。
SLC24A5遺伝子の変化こそ「白人」を生みだしたと言えるものだからね。
ただ、これも碧眼と同様、7000〜10000年前に誕生したらしい。
さらに新しく5300〜6000年前という説もある。
だから、金髪・碧眼・白い肌という人種的特徴と印欧語族の拡大とがリンクしているのは確実だね。
さらに、遺伝子のR1aとR1bがこれに重なる。
コーカソイドのひとつの核がこの連中だろうね。
これは最新の論文かな。
よく読んでみよう。
SLC24A5遺伝子の変化こそ「白人」を生みだしたと言えるものだからね。
ただ、これも碧眼と同様、7000〜10000年前に誕生したらしい。
さらに新しく5300〜6000年前という説もある。
だから、金髪・碧眼・白い肌という人種的特徴と印欧語族の拡大とがリンクしているのは確実だね。
さらに、遺伝子のR1aとR1bがこれに重なる。
コーカソイドのひとつの核がこの連中だろうね。
>>573
ちっと古いから前のやつかな
Reconstructing Indian Population History
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2842210/
と思ったがこれかな
Genetic evidence for recent population mixture in India.
http://www.ncbi.nlm.nih.gov/pubmed/23932107
http://www.cell.com/AJHG/retrieve/pii/S0002929713003248
http://www.sciencedirect.com/science/article/pii/S0002929713003248
ちっと古いから前のやつかな
Reconstructing Indian Population History
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2842210/
と思ったがこれかな
Genetic evidence for recent population mixture in India.
http://www.ncbi.nlm.nih.gov/pubmed/23932107
http://www.cell.com/AJHG/retrieve/pii/S0002929713003248
http://www.sciencedirect.com/science/article/pii/S0002929713003248
>>610
書けたがパスww
>>574
この辺りやろう
Complete Mitochondrial Genomes of Ancient Canids Suggest a European Origin of Domestic Dogs
http://www.sciencemag.org/content/342/6160/871.abstract
【生物】犬の起源は欧州
http://anago.2ch.net/test/read.cgi/scienceplus/1384550380/1
書けたがパスww
>>574
この辺りやろう
Complete Mitochondrial Genomes of Ancient Canids Suggest a European Origin of Domestic Dogs
http://www.sciencemag.org/content/342/6160/871.abstract
【生物】犬の起源は欧州
http://anago.2ch.net/test/read.cgi/scienceplus/1384550380/1
イスラエルに棲息している野生のイノシシの起源はヨーロッパ、古代人がヨーロッパから持ち込んだ可能性
http://jp.sciencenewsline.com/articles/2013110423390003.html
イスラエルに棲息している野生のイノシシは世界中に棲息している他のイノシシと似ている。
そのため、これまでイスラエルに棲息している野生のイノシシはエジプトやイランから、
中東へと広がった他のイノシシの親戚だと考えられてきたがそうした考えは間違いであることが判った。
Tel Aviv Universityの考古学研究チームと古代中東文化研究チームが共同研究の結果から、
イスラエルに棲息している野生のイノシシはイスラエルの周囲の国々に棲息しているイノシシとは異なり、
ヨーロッパに起源をもつことを発見した。
遺伝子解析と考古学的分析の手法を用いることにより、
研究チームは、イスラエルに棲息している野生のイノシシは3000年前に
ペリシテ人や他の侵略者によってイスラエルに持ち込まれた家畜のブタが野生化したものだという可能性を見つけた。
そして研究チームはこの研究成果を今週号の「Scientific Reports」を通じて発表した。
「我々が行った遺伝子解析の結果から、
イスラエルに棲息している野生のイノシシは紀元前900年頃の鉄器時代のヨーロッパで飼育されていたブタの子孫であることが証明されたのです」
とTel Aviv UniversityのProf. Finkelsteinは述べる。
「また、ペリシテ人の遺跡から大量のブタの骨が見つかっていることを考慮すると、ヨーロッパのブタは、
ペリシテ人の船舶によってイスラエルに運ばれてきたものだということが言えるかもしれません。」
イスラエル南部の海岸沿いにあるペリシテ人の遺跡で見つかってきたブタの骨は、紀元前1150〜950年頃の鉄器時代の遡ることができるが、
こうしたブタの骨は、イスラエルの同年代の他遺跡ではまったく見つからないか、見つかったとしても非常に稀な存在となる。
http://jp.sciencenewsline.com/articles/2013110423390003.html
イスラエルに棲息している野生のイノシシは世界中に棲息している他のイノシシと似ている。
そのため、これまでイスラエルに棲息している野生のイノシシはエジプトやイランから、
中東へと広がった他のイノシシの親戚だと考えられてきたがそうした考えは間違いであることが判った。
Tel Aviv Universityの考古学研究チームと古代中東文化研究チームが共同研究の結果から、
イスラエルに棲息している野生のイノシシはイスラエルの周囲の国々に棲息しているイノシシとは異なり、
ヨーロッパに起源をもつことを発見した。
遺伝子解析と考古学的分析の手法を用いることにより、
研究チームは、イスラエルに棲息している野生のイノシシは3000年前に
ペリシテ人や他の侵略者によってイスラエルに持ち込まれた家畜のブタが野生化したものだという可能性を見つけた。
そして研究チームはこの研究成果を今週号の「Scientific Reports」を通じて発表した。
「我々が行った遺伝子解析の結果から、
イスラエルに棲息している野生のイノシシは紀元前900年頃の鉄器時代のヨーロッパで飼育されていたブタの子孫であることが証明されたのです」
とTel Aviv UniversityのProf. Finkelsteinは述べる。
「また、ペリシテ人の遺跡から大量のブタの骨が見つかっていることを考慮すると、ヨーロッパのブタは、
ペリシテ人の船舶によってイスラエルに運ばれてきたものだということが言えるかもしれません。」
イスラエル南部の海岸沿いにあるペリシテ人の遺跡で見つかってきたブタの骨は、紀元前1150〜950年頃の鉄器時代の遡ることができるが、
こうしたブタの骨は、イスラエルの同年代の他遺跡ではまったく見つからないか、見つかったとしても非常に稀な存在となる。
615 :世界@名無史さん:2013/12/22(日) 08:09:09.28 0
印欧語族が連れてきたのかな。
>>614
旧約聖書のペリシテ人の姿を解き明かす 古代都市ガトの発掘調査
http://www.archeonews.net/2011/07/blog-post_11.html
約3000年前に、ペリシテ人は地中海岸周辺に居住しており、ガトは新たに開拓された都市だった。
この都市で最も有名な住民は、サムエル記にある羊飼いの若者ダビデの投石によって倒された巨人ゴリアテである。
(略)
遺跡からは古代ギリシア様式の文様がある土器片が発見されており、ペリシテ人の起源がエーゲ海方面にあることを示唆している。
ペリシテ人は現在のギリシア地域から紀元前1200年ごろに海を渡って到来した。
そしてアシュケロンやアシュドドなどの主要な港、イスラエルの都市群、現在のパレスチナの領域であるガザ地区などを支配するようになった。
ガトの遺跡の発掘によって、ペリシテ人達は在地の文化を一部受け入れつつも、自分たちのルーツを忘れていなかったことがわかっている。
例えば、この地域に到来してから5世紀たった後でも、彼らはギリシア語の名前を持つ神々を崇めていた。
調査によって、ペリシテ人はエーゲ海地域と同様、レンズ豆を多く摂取していたことがわかっている。
遺跡で廃棄されていた動物骨の調査から、彼らは豚と犬を食べていたようである。これらはイスラエル人とは異なっており、イスラエル人はこれらの動物を不浄なものとみなしていた。この戒律は現在のユダヤ教にも残っている。
また、ガトの町が紀元前9世紀に破壊を受けたことも発見された。
市の周囲には包囲群によって作られた溝や盛り土が出土しており、現在でも都市があった丘陵部の周りに黒いラインとなって見えている。
この時期に起きたガトの壊滅はアラム人の王ハザエルによって紀元前830年に行われたもののようである。
この出来事は旧約聖書の列王記に言及されている。
このように、ガトは紀元前10世紀から前9世紀にかけてのペリシテ人の生活の様子を明らかにする、極めて豊富な物的証拠を提供してくれている。
その中にはイェルサレムのダビデ王やソロモン王によって統治されていた時代も含まれる。
他のペリシテ人の町の遺跡は、その前後の時代の情報は提供しているが、この重要な時代の資料がない。
ガトは、そのギャップを埋めてくれているのだ。
旧約聖書のペリシテ人の姿を解き明かす 古代都市ガトの発掘調査
http://www.archeonews.net/2011/07/blog-post_11.html
約3000年前に、ペリシテ人は地中海岸周辺に居住しており、ガトは新たに開拓された都市だった。
この都市で最も有名な住民は、サムエル記にある羊飼いの若者ダビデの投石によって倒された巨人ゴリアテである。
(略)
遺跡からは古代ギリシア様式の文様がある土器片が発見されており、ペリシテ人の起源がエーゲ海方面にあることを示唆している。
ペリシテ人は現在のギリシア地域から紀元前1200年ごろに海を渡って到来した。
そしてアシュケロンやアシュドドなどの主要な港、イスラエルの都市群、現在のパレスチナの領域であるガザ地区などを支配するようになった。
ガトの遺跡の発掘によって、ペリシテ人達は在地の文化を一部受け入れつつも、自分たちのルーツを忘れていなかったことがわかっている。
例えば、この地域に到来してから5世紀たった後でも、彼らはギリシア語の名前を持つ神々を崇めていた。
調査によって、ペリシテ人はエーゲ海地域と同様、レンズ豆を多く摂取していたことがわかっている。
遺跡で廃棄されていた動物骨の調査から、彼らは豚と犬を食べていたようである。これらはイスラエル人とは異なっており、イスラエル人はこれらの動物を不浄なものとみなしていた。この戒律は現在のユダヤ教にも残っている。
また、ガトの町が紀元前9世紀に破壊を受けたことも発見された。
市の周囲には包囲群によって作られた溝や盛り土が出土しており、現在でも都市があった丘陵部の周りに黒いラインとなって見えている。
この時期に起きたガトの壊滅はアラム人の王ハザエルによって紀元前830年に行われたもののようである。
この出来事は旧約聖書の列王記に言及されている。
このように、ガトは紀元前10世紀から前9世紀にかけてのペリシテ人の生活の様子を明らかにする、極めて豊富な物的証拠を提供してくれている。
その中にはイェルサレムのダビデ王やソロモン王によって統治されていた時代も含まれる。
他のペリシテ人の町の遺跡は、その前後の時代の情報は提供しているが、この重要な時代の資料がない。
ガトは、そのギャップを埋めてくれているのだ。
617 :世界@名無史さん:2013/12/27(金) 00:12:50.82 0
エサウのモデルも気になる
氷河期にヨーロッパ人が大西洋を渡っていたという新説
http://www.archeonews.net/2012/03/blog-post_12.html
最初にアメリカ大陸へ渡ったのはアジア系の人々であり、彼らがいわゆるネイティブ・アメリカンの祖となったというのが通説だ。
しかし、アジア人よりも1万年も前に、ヨーロッパに起源を持つ人々が北大西洋を渡ってアメリカ大陸に到達していたという新説が発表された。
氷河期の北半球 灰色になっているのが氷河期の最盛期に氷に覆われていたと考えられる範囲
2万6000年前〜1万9000年に年代づけられる、ヨーロッパの様式と類似した石器が、アメリカ東海岸沿いの6地点で発見されていた。
その内3つはメリーランドのデルマーバ半島に位置しており、デラウェア大学の考古学者Darrin Lowery博士によって発見された。
あと1つはペンシルヴァニア、もう1つはヴァージニアである。
最後はホタテ貝の底引き網漁によってヴァージニア沖60マイルの海底から発見された。氷河期にはこの場所は陸地だった。
ヨーロッパのソリュトレアン文化の石器とアメリカ東海岸の遺跡で過去に発見された石器が似ていることはこれまでにも指摘されていた。
ソリュトレアン文化はフランスやイベリア半島で発見されている。
しかし、これまでにアメリカ東海岸で発見された石器は約1万5000年前のものであり、当時のヨーロッパではすでに作られなくなっていたタイプのものだ。
そのため、ほとんどの考古学者はこれらの石器に関連があると考えていなかった。
しかし、新たに発見された遺跡の石器は、2万6000年から1万9000年前のものであることが分かった。
この年代は、西ヨーロッパのソリュトレアン文化とほぼ同時代にあたる。
さらに、1971年にヴァージニアで発見されたヨーロッパのスタイルの石器に対する理化学的な分析が、昨年行われた。
その結果、石材がフランスで採取されたものだということが判明したのである。
アメリカ・スミソニアン研究所のDennis Stanford教授と、イギリス・エクセター大学のBruce Bradley教授が網羅的な分析を行った。
結果として、西ヨーロッパの石器時代の人々が、氷河期の真っただ中に大西洋北辺の凍りついた場所を通って、北米に移住していたという説を唱えるにいたったのである。
http://www.archeonews.net/2012/03/blog-post_12.html
最初にアメリカ大陸へ渡ったのはアジア系の人々であり、彼らがいわゆるネイティブ・アメリカンの祖となったというのが通説だ。
しかし、アジア人よりも1万年も前に、ヨーロッパに起源を持つ人々が北大西洋を渡ってアメリカ大陸に到達していたという新説が発表された。
氷河期の北半球 灰色になっているのが氷河期の最盛期に氷に覆われていたと考えられる範囲
2万6000年前〜1万9000年に年代づけられる、ヨーロッパの様式と類似した石器が、アメリカ東海岸沿いの6地点で発見されていた。
その内3つはメリーランドのデルマーバ半島に位置しており、デラウェア大学の考古学者Darrin Lowery博士によって発見された。
あと1つはペンシルヴァニア、もう1つはヴァージニアである。
最後はホタテ貝の底引き網漁によってヴァージニア沖60マイルの海底から発見された。氷河期にはこの場所は陸地だった。
ヨーロッパのソリュトレアン文化の石器とアメリカ東海岸の遺跡で過去に発見された石器が似ていることはこれまでにも指摘されていた。
ソリュトレアン文化はフランスやイベリア半島で発見されている。
しかし、これまでにアメリカ東海岸で発見された石器は約1万5000年前のものであり、当時のヨーロッパではすでに作られなくなっていたタイプのものだ。
そのため、ほとんどの考古学者はこれらの石器に関連があると考えていなかった。
しかし、新たに発見された遺跡の石器は、2万6000年から1万9000年前のものであることが分かった。
この年代は、西ヨーロッパのソリュトレアン文化とほぼ同時代にあたる。
さらに、1971年にヴァージニアで発見されたヨーロッパのスタイルの石器に対する理化学的な分析が、昨年行われた。
その結果、石材がフランスで採取されたものだということが判明したのである。
アメリカ・スミソニアン研究所のDennis Stanford教授と、イギリス・エクセター大学のBruce Bradley教授が網羅的な分析を行った。
結果として、西ヨーロッパの石器時代の人々が、氷河期の真っただ中に大西洋北辺の凍りついた場所を通って、北米に移住していたという説を唱えるにいたったのである。
>>618
http://2.bp.blogspot.com/-uz2kYLctldE/T12N4z7jt8I/AAAAAAAABX0/Aa7g1z9-fsM/s1600/600px-Iceage_north-intergl_glac_hg.png
http://2.bp.blogspot.com/-uz2kYLctldE/T12N4z7jt8I/AAAAAAAABX0/Aa7g1z9-fsM/s1600/600px-Iceage_north-intergl_glac_hg.png
{kind=link}
>>619
氷河期のピークには、北大西洋の300万平方マイルが、1年中もしくは季節的に厚い氷に覆われていた。
しかし、季節的に氷が溶け、海に面していたところは、アザラシや海鳥、魚、今は絶命している北極ペンギンに似た種類の動物、オオウミガラスなど、豊富な食料資源があった。
Stanford教授とBradley教授は、石器時代の人々が大西洋の全長1500マイルの氷の上を移動する能力があったと過去にも述べていたが、この説を裏付ける証拠がほとんどなかった。
しかし、アメリカ東海岸で発見された石器とその理化学的分析の結果は、状況を変えつつある。
これから、テネシー、メリーランド、テキサスなどで新たに調査がスタートする予定だ。
調査によってさらなる証拠をもたらしてくれるかもしれない。
Stanford教授とBradley教授の説を裏付けるもう一つの事実は、シベリア北東部やアラスカで1万5500年よりも前の時代の人類による活動の痕跡が発見されていないことだ。
もしアメリカ東海岸で発見された石器を持っていた人々が、ヨーロッパではなくアジアから来ていたのであれば、これらの2つの地域で1万9000年よりも前の活動の痕跡があるはずだ。
アジアからの人々は1万5500年前にベーリング海峡もしくはアリューシャン列島を通ってアメリカ大陸にやってきた。
この後は気候もずっと人間には適しており、ヨーロッパ人よりも多くの数のアジア人が移住することができたと考えられる。
ヨーロッパ起源のネイティブ・アメリカンは新しく来たアジア系の人々に吸収されてしまったか、物理的に全滅させられたもしくは食料の獲得競争に敗れて絶滅してしまった。
西ヨーロッパの石器時代の人々に起源を持つ遺伝子は北東アジアには存在しない。
しかし北米のネイティブ・アメリカンの集団には、わずかな量であるが遺伝子が見られるのだ。
フロリダで発見された8000年前の人骨から採取されたDNAの分析から、ヨーロッパに起源を持つ可能性がある遺伝的特徴が発見されている。
また、他とは独立して存在していたごくわずかなネイティブ・アメリカンの中には、アジア起源のネイティブ・アメリカンとは異なる言語を使用していた。
ホタテ貝の底引き網漁によって発見された遺跡は、今夏、詳細な調査が行われる予定だ。
(略)この板は短すぎるな
氷河期のピークには、北大西洋の300万平方マイルが、1年中もしくは季節的に厚い氷に覆われていた。
しかし、季節的に氷が溶け、海に面していたところは、アザラシや海鳥、魚、今は絶命している北極ペンギンに似た種類の動物、オオウミガラスなど、豊富な食料資源があった。
Stanford教授とBradley教授は、石器時代の人々が大西洋の全長1500マイルの氷の上を移動する能力があったと過去にも述べていたが、この説を裏付ける証拠がほとんどなかった。
しかし、アメリカ東海岸で発見された石器とその理化学的分析の結果は、状況を変えつつある。
これから、テネシー、メリーランド、テキサスなどで新たに調査がスタートする予定だ。
調査によってさらなる証拠をもたらしてくれるかもしれない。
Stanford教授とBradley教授の説を裏付けるもう一つの事実は、シベリア北東部やアラスカで1万5500年よりも前の時代の人類による活動の痕跡が発見されていないことだ。
もしアメリカ東海岸で発見された石器を持っていた人々が、ヨーロッパではなくアジアから来ていたのであれば、これらの2つの地域で1万9000年よりも前の活動の痕跡があるはずだ。
アジアからの人々は1万5500年前にベーリング海峡もしくはアリューシャン列島を通ってアメリカ大陸にやってきた。
この後は気候もずっと人間には適しており、ヨーロッパ人よりも多くの数のアジア人が移住することができたと考えられる。
ヨーロッパ起源のネイティブ・アメリカンは新しく来たアジア系の人々に吸収されてしまったか、物理的に全滅させられたもしくは食料の獲得競争に敗れて絶滅してしまった。
西ヨーロッパの石器時代の人々に起源を持つ遺伝子は北東アジアには存在しない。
しかし北米のネイティブ・アメリカンの集団には、わずかな量であるが遺伝子が見られるのだ。
フロリダで発見された8000年前の人骨から採取されたDNAの分析から、ヨーロッパに起源を持つ可能性がある遺伝的特徴が発見されている。
また、他とは独立して存在していたごくわずかなネイティブ・アメリカンの中には、アジア起源のネイティブ・アメリカンとは異なる言語を使用していた。
ホタテ貝の底引き網漁によって発見された遺跡は、今夏、詳細な調査が行われる予定だ。
(略)この板は短すぎるな
>>574
イギリス人の祖先は何者か? DNAによる人の移動の分析
http://www.archeonews.net/2011/09/dna.html
オックスフォード大学とエジンバラ大学の研究者らによると、ヨーロッパの男性の大半はそれよりもずっと前の段階にヨーロッパに定住していた人々(おそらくは狩猟採集民)と遺伝的なつながりがあるという。
この研究成果はヨーロッパ人男性に共通する遺伝子に基づいており、昨年報告された分析結果を検証する目的があった。
昨年の報告では、イギリス人は近東からの農耕の拡大ととともに西へ移動してきた人々の子孫であるとしていた。
1億人のヨーロッパ人がR-M269と呼ばれる遺伝子を有しており、その中にはイギリス人の4分の3も含まれる。
ヨーロッパにおける人の動きを理解する鍵は、この遺伝子を持つグループがいつヨーロッパに拡散していったか、を明らかにすることである。
研究者らによると、過去の分析で行ったこのグループの遺伝子による年代推定の結果は、極めて多種多様であった。
また、過去に報告していた東から西の分布傾向は、より広範で包括的なデータ・セットからは確認することができなかったとしている。
したがって、農耕民による人口の拡散に関する証拠は見られない、と両大学の研究者らは結論付けている。
オックスフォード大学の遺伝学者でこの研究を主導しているCristian Capelli博士によれば、新しい研究の成果がこれまでのヨーロッパにおける人口の動きに関する議論を「リセット」するものだという。
「我々の研究は、ヨーロッパ人のY染色体が農耕民によってもたらされたとする近年の研究をひっくり返す結果となった」と博士は述べている。
共同研究者でエジンバラ大学Jim Wilson博士は、この研究によって遺伝子の分析が年代推定の正確さに大きな影響を与えた初めての例になる、と付け加えている。
「祖先の系統が発生した時期を推定することは、遺伝学の興味深い応用分野だが、正確な年代を得ることが非常に難しい。多くの人は遺伝子に関する資料が増えれば年代もより正確になると考えているようだが、そうではない。
遺伝子にある種の目印があることの方が、より年代推定には最も適しているのだ」
この研究ではまた、ヨーロッパの各地で共通するR-M269の下位グループ群が、各地で拡散していく様子も報告されている。
イギリス人の祖先は何者か? DNAによる人の移動の分析
http://www.archeonews.net/2011/09/dna.html
オックスフォード大学とエジンバラ大学の研究者らによると、ヨーロッパの男性の大半はそれよりもずっと前の段階にヨーロッパに定住していた人々(おそらくは狩猟採集民)と遺伝的なつながりがあるという。
この研究成果はヨーロッパ人男性に共通する遺伝子に基づいており、昨年報告された分析結果を検証する目的があった。
昨年の報告では、イギリス人は近東からの農耕の拡大ととともに西へ移動してきた人々の子孫であるとしていた。
1億人のヨーロッパ人がR-M269と呼ばれる遺伝子を有しており、その中にはイギリス人の4分の3も含まれる。
ヨーロッパにおける人の動きを理解する鍵は、この遺伝子を持つグループがいつヨーロッパに拡散していったか、を明らかにすることである。
研究者らによると、過去の分析で行ったこのグループの遺伝子による年代推定の結果は、極めて多種多様であった。
また、過去に報告していた東から西の分布傾向は、より広範で包括的なデータ・セットからは確認することができなかったとしている。
したがって、農耕民による人口の拡散に関する証拠は見られない、と両大学の研究者らは結論付けている。
オックスフォード大学の遺伝学者でこの研究を主導しているCristian Capelli博士によれば、新しい研究の成果がこれまでのヨーロッパにおける人口の動きに関する議論を「リセット」するものだという。
「我々の研究は、ヨーロッパ人のY染色体が農耕民によってもたらされたとする近年の研究をひっくり返す結果となった」と博士は述べている。
共同研究者でエジンバラ大学Jim Wilson博士は、この研究によって遺伝子の分析が年代推定の正確さに大きな影響を与えた初めての例になる、と付け加えている。
「祖先の系統が発生した時期を推定することは、遺伝学の興味深い応用分野だが、正確な年代を得ることが非常に難しい。多くの人は遺伝子に関する資料が増えれば年代もより正確になると考えているようだが、そうではない。
遺伝子にある種の目印があることの方が、より年代推定には最も適しているのだ」
この研究ではまた、ヨーロッパの各地で共通するR-M269の下位グループ群が、各地で拡散していく様子も報告されている。
622 :世界@名無史さん:2013/12/27(金) 03:43:10.15 0
>>621
古い!
古い!
>ヨーロッパ起源のネイティブ・アメリカンは新しく来たアジア系の人々に吸収されてしまったか、
物理的に全滅させられたもしくは食料の獲得競争に敗れて絶滅してしまった。
つまり元々新大陸は白人のものだったんだな
モンゴロイドに奪われた大陸を取り返しただけだったんだ
物理的に全滅させられたもしくは食料の獲得競争に敗れて絶滅してしまった。
つまり元々新大陸は白人のものだったんだな
モンゴロイドに奪われた大陸を取り返しただけだったんだ
624 :世界@名無史さん:2013/12/27(金) 16:58:25.64 0
ということにしたいんだろうなあ
625 :世界@名無史さん:2013/12/27(金) 22:05:09.55 0
皮肉にも白人が滅ぼしたベオスック(Beothuk)族に、このヨーロッパ起源説を解く鍵が隠されてたのかもな
626 :世界@名無史さん:2013/12/27(金) 22:12:26.95 0
容姿端麗かどうかは素顔を見てから判断したい。
目の周りの化粧が凄くて。
目の周りの化粧が凄くて。
ライオンマン
http://ja.wikipedia.org/wiki/%E3%83%A9%E3%82%A4%E3%82%AA%E3%83%B3%E3%83%9E%E3%83%B3
概要[編集]
この獅子頭の象牙彫刻は、まずライオンマン(独: Lowenmensch、直訳すると "ライオン人")、
次いでライオンレディ(独: Lowenfrau)と呼ばれた。これは世界最古の動物形象の彫刻であると同時に、
いわゆる彫刻として知られる最古のもののひとつである。
この彫刻は、動物に人間の性質を擬した擬人化であると解釈されているが、
神の表現である可能性もある。発見されたのと同じ地層の放射性炭素年代測定により、この小立像は約32,000年前のものとされている[1][2]。
これは考古学上、オーリニャック文化のものとされている[3]。
来歴[編集]
この像の断片は1939年に、ドイツのシュヴァーベン・アルプ(英語版)のローネタール(はぐれ谷)にある
ホーレンシュタイン・シュターデル (en) の洞窟(ホーレンシュタイン山のシュターデル洞窟)で見つかった。
しかし第二次世界大戦が始まったため、それは忘れ去られ、再発見されたのは30年後だった。
最初の修復によって、頭部の無い人型の小立像であることが分かった。1997-1998年に彫刻の残りの断片が見つかり、頭部が再構築され取り付けられた。
特徴[編集]
彫刻は高さ29.6センチ、幅5.6センチ、奥行5.9センチである。
燧石でできた石製ナイフを使い、マンモスの象牙から刻み出された。
左腕に、横方向の曲がった線が7本平行に刻まれている。
動物的な特徴としては、ライオンの頭部、すらりと長い体躯、獣の後肢のような腕が挙げられる。
人間的な特徴としては、人間的な脚部と足、直立して伸びた背筋が挙げられる[4]。
http://ja.wikipedia.org/wiki/%E3%83%95%E3%82%A1%E3%82%A4%E3%83%AB:Lion_man_photo.jpg
http://ja.wikipedia.org/wiki/%E3%83%A9%E3%82%A4%E3%82%AA%E3%83%B3%E3%83%9E%E3%83%B3
概要[編集]
この獅子頭の象牙彫刻は、まずライオンマン(独: Lowenmensch、直訳すると "ライオン人")、
次いでライオンレディ(独: Lowenfrau)と呼ばれた。これは世界最古の動物形象の彫刻であると同時に、
いわゆる彫刻として知られる最古のもののひとつである。
この彫刻は、動物に人間の性質を擬した擬人化であると解釈されているが、
神の表現である可能性もある。発見されたのと同じ地層の放射性炭素年代測定により、この小立像は約32,000年前のものとされている[1][2]。
これは考古学上、オーリニャック文化のものとされている[3]。
来歴[編集]
この像の断片は1939年に、ドイツのシュヴァーベン・アルプ(英語版)のローネタール(はぐれ谷)にある
ホーレンシュタイン・シュターデル (en) の洞窟(ホーレンシュタイン山のシュターデル洞窟)で見つかった。
しかし第二次世界大戦が始まったため、それは忘れ去られ、再発見されたのは30年後だった。
最初の修復によって、頭部の無い人型の小立像であることが分かった。1997-1998年に彫刻の残りの断片が見つかり、頭部が再構築され取り付けられた。
特徴[編集]
彫刻は高さ29.6センチ、幅5.6センチ、奥行5.9センチである。
燧石でできた石製ナイフを使い、マンモスの象牙から刻み出された。
左腕に、横方向の曲がった線が7本平行に刻まれている。
動物的な特徴としては、ライオンの頭部、すらりと長い体躯、獣の後肢のような腕が挙げられる。
人間的な特徴としては、人間的な脚部と足、直立して伸びた背筋が挙げられる[4]。
http://ja.wikipedia.org/wiki/%E3%83%95%E3%82%A1%E3%82%A4%E3%83%AB:Lion_man_photo.jpg
{kind=link}
これじゃない
ヴィレンドルフのヴィーナス
http://en.wikipedia.org/wiki/Venus_of_Willendorf
Dating[edit]
Venus of WillendorfAfter a wide variety of proposed dates, following a revised analysis of the stratigraphy of its site in 1990,
the figure was estimated to have been carved 24,000?22,000 BCE,[4]
but more recent estimates have pushed the date back slightly to between about 28,000 and 25,000 BCE.
It is believed that the figure was carved during the Paleolithic Period,
also known as the "Old Stone Age". This period of History started around 30,000 BCE.[6]
http://en.wikipedia.org/wiki/File:Venus_von_Willendorf_01.jpg
http://en.wikipedia.org/wiki/File:Willendorf-Venus-1468.jpg
ヴィレンドルフのヴィーナス
http://en.wikipedia.org/wiki/Venus_of_Willendorf
Dating[edit]
Venus of WillendorfAfter a wide variety of proposed dates, following a revised analysis of the stratigraphy of its site in 1990,
the figure was estimated to have been carved 24,000?22,000 BCE,[4]
but more recent estimates have pushed the date back slightly to between about 28,000 and 25,000 BCE.
It is believed that the figure was carved during the Paleolithic Period,
also known as the "Old Stone Age". This period of History started around 30,000 BCE.[6]
http://en.wikipedia.org/wiki/File:Venus_von_Willendorf_01.jpg
{kind=link}
http://en.wikipedia.org/wiki/File:Willendorf-Venus-1468.jpg
{kind=link}
>>629
一つはこれ、もう一つが思い出せない、、、
ホーレ・フェルスのヴィーナス
http://ja.wikipedia.org/wiki/%E3%83%9B%E3%83%BC%E3%83%AC%E3%83%BB%E3%83%95%E3%82%A7%E3%83%AB%E3%82%B9%E3%81%AE%E3%83%B4%E3%82%A3%E3%83%BC%E3%83%8A%E3%82%B9
ホーレ・フェルスのヴィーナス (実際の大きさは高さ6cm、幅が3cm、重さが33g程度[1])
ヴィーナス像が発見されたホーレ・フェルスの洞窟内部ホーレ・フェルスのヴィーナス (Venus vom Hohlen Fels, vom Hohle Fels)は
2008年にドイツのシェルクリンゲン近郊で発見された後期旧石器時代のヴィーナス像。
この時代はじめの初期オーリニャック文化でつくられたと考えられ、年代的にはおよそ3万5千年から4万年ほど前にあたる。
描かれているのはおそらくヨーロッパにかつて分布した新人であるクロマニヨン人の姿である。
疑いようもなく旧石器時代では最古の美術品であり、あるいはより一般的な先史時代の造形美術としても最も古い。
テュービンゲン大学では2011年の段階でもこの小像の調査が続いているが、
いずれこの地域から出土した他の発掘品もふくめてシュヴァーベンに新たに建設予定の博物館におさめられる予定である[2]。
http://upload.wikimedia.org/wikipedia/commons/1/18/VenusHohlefels2.jpg
>>628
http://upload.wikimedia.org/wikipedia/commons/f/f8/Lion_man_photo.jpg
>>629
http://upload.wikimedia.org/wikipedia/commons/5/50/Venus_von_Willendorf_01.jpg
http://upload.wikimedia.org/wikipedia/commons/f/f7/Willendorf-Venus-1468.jpg
一つはこれ、もう一つが思い出せない、、、
ホーレ・フェルスのヴィーナス
http://ja.wikipedia.org/wiki/%E3%83%9B%E3%83%BC%E3%83%AC%E3%83%BB%E3%83%95%E3%82%A7%E3%83%AB%E3%82%B9%E3%81%AE%E3%83%B4%E3%82%A3%E3%83%BC%E3%83%8A%E3%82%B9
ホーレ・フェルスのヴィーナス (実際の大きさは高さ6cm、幅が3cm、重さが33g程度[1])
ヴィーナス像が発見されたホーレ・フェルスの洞窟内部ホーレ・フェルスのヴィーナス (Venus vom Hohlen Fels, vom Hohle Fels)は
2008年にドイツのシェルクリンゲン近郊で発見された後期旧石器時代のヴィーナス像。
この時代はじめの初期オーリニャック文化でつくられたと考えられ、年代的にはおよそ3万5千年から4万年ほど前にあたる。
描かれているのはおそらくヨーロッパにかつて分布した新人であるクロマニヨン人の姿である。
疑いようもなく旧石器時代では最古の美術品であり、あるいはより一般的な先史時代の造形美術としても最も古い。
テュービンゲン大学では2011年の段階でもこの小像の調査が続いているが、
いずれこの地域から出土した他の発掘品もふくめてシュヴァーベンに新たに建設予定の博物館におさめられる予定である[2]。
http://upload.wikimedia.org/wikipedia/commons/1/18/VenusHohlefels2.jpg
{kind=link}
>>628
http://upload.wikimedia.org/wikipedia/commons/f/f8/Lion_man_photo.jpg
{kind=link}
>>629
http://upload.wikimedia.org/wikipedia/commons/5/50/Venus_von_Willendorf_01.jpg
{kind=link}
http://upload.wikimedia.org/wikipedia/commons/f/f7/Willendorf-Venus-1468.jpg
{kind=link}
http://oka-jp.seesaa.net/article/317393091.html
http://jomontaro.web.fc2.com/newpage176.html
http://en.wikipedia.org/wiki/Venus_figurines
ここに色々あった
探していたのはこれ
http://en.wikipedia.org/wiki/Venus_of_Lespugue
http://upload.wikimedia.org/wikipedia/commons/8/89/Venus_de_Lespugue_%28replica%29.jpg
http://donsmaps.com/lespuguevenus.html
The Venus of Lespugue
is a Venus figurine, a statuette of a nude female figure of the Gravettian, dated to between 26,000 and 24,000 years ago.
http://jomontaro.web.fc2.com/newpage176.html
http://en.wikipedia.org/wiki/Venus_figurines
ここに色々あった
探していたのはこれ
http://en.wikipedia.org/wiki/Venus_of_Lespugue
http://upload.wikimedia.org/wikipedia/commons/8/89/Venus_de_Lespugue_%28replica%29.jpg
{kind=link}
http://donsmaps.com/lespuguevenus.html
The Venus of Lespugue
is a Venus figurine, a statuette of a nude female figure of the Gravettian, dated to between 26,000 and 24,000 years ago.
>>548
古代フェニキア人のDNA、現代人にも
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=89591997
タイラースミス氏らは、DNAサンプルと歴史的、考古学的な記録とを照合しながら研究を進めた。
かつてフェニキア人の交易の中心であった地中海諸国(シリア、パレスチナ、チュニジア、モロッコ、キプロス、マルタ)の男性1330人からY染色体が採取され、
分析が行われた。
母系で受け継がれるミトコンドリアDNAとは異なり、父系で継承されるY染色体からはより詳細な遺伝情報が得られると考えられている。
Y染色体のデータを分析した結果、地中海周辺地域にはフェニキア人に関連する遺伝子系統が少なくとも7系統あることが明らかになった。
フェニキア人が交易先の各地に遺伝子を広め、いまも該当地域の6%の人に継承されている可能性が示唆された。
「フェニキア人の名残は現代の人々にも受け継がれていることが示された」とタイラースミス氏は話す。
この研究は「American Journal of Human Genetics」誌の最新号に掲載されている。
古代フェニキア人のDNA、現代人にも
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=89591997
タイラースミス氏らは、DNAサンプルと歴史的、考古学的な記録とを照合しながら研究を進めた。
かつてフェニキア人の交易の中心であった地中海諸国(シリア、パレスチナ、チュニジア、モロッコ、キプロス、マルタ)の男性1330人からY染色体が採取され、
分析が行われた。
母系で受け継がれるミトコンドリアDNAとは異なり、父系で継承されるY染色体からはより詳細な遺伝情報が得られると考えられている。
Y染色体のデータを分析した結果、地中海周辺地域にはフェニキア人に関連する遺伝子系統が少なくとも7系統あることが明らかになった。
フェニキア人が交易先の各地に遺伝子を広め、いまも該当地域の6%の人に継承されている可能性が示唆された。
「フェニキア人の名残は現代の人々にも受け継がれていることが示された」とタイラースミス氏は話す。
この研究は「American Journal of Human Genetics」誌の最新号に掲載されている。
>>633
"Phoenician" Y-chromosomes
http://dienekes.blogspot.jp/2008/10/phoenician-y-chromosomes.html
http://2.bp.blogspot.com/_ro2ijOk8JWc/SQoI4CPBFMI/AAAAAAAAAak/GFXiy3UvQB0/s1600-h/phoenicians.jpg
Identifying Genetic Traces of Historical Expansions: Phoenician Footprints in the Mediterranean
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2668035/
"Phoenician" Y-chromosomes
http://dienekes.blogspot.jp/2008/10/phoenician-y-chromosomes.html
http://2.bp.blogspot.com/_ro2ijOk8JWc/SQoI4CPBFMI/AAAAAAAAAak/GFXiy3UvQB0/s1600-h/phoenicians.jpg
{kind=link}
Identifying Genetic Traces of Historical Expansions: Phoenician Footprints in the Mediterranean
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2668035/
635 :世界@名無史さん:2013/12/29(日) 15:44:55.66 0
>>633 ナポレオン一族もそうなのかな。E系だけど
636 :世界@名無史さん:2013/12/29(日) 15:48:44.38 0
白人って寒い所済んでるのに凹凸が大きいのは何故だろう
637 :世界@名無史さん:2013/12/29(日) 19:44:35.65 0
酷寒地域には住んでない。
638 :世界@名無史さん:2013/12/30(月) 21:31:50.98 0
最寒月平均気温-30℃以下の土地で平たい顔族は生まれたということかな
そんな土地は氷河期には、ヨーロッパにも中東にも人が住める場所にはなかった
そんな土地は氷河期には、ヨーロッパにも中東にも人が住める場所にはなかった
639 :世界@名無史さん:2014/01/03(金) 10:28:05.72 0
シベリア(モンゴル高原含む)は世界の寒極だから、
表面積を極小化した人種が誕生したのだろうね。
みなみにアルプス以北の欧州はヴュルム氷期の大半は無人地帯だった。
表面積を極小化した人種が誕生したのだろうね。
みなみにアルプス以北の欧州はヴュルム氷期の大半は無人地帯だった。
海流を変えれらたら欧州消滅できるかも
641 :世界@名無史さん:2014/01/03(金) 15:30:54.02 0
地中海地域に北方から環境難民が殺到
642 :世界@名無史さん:2014/01/04(土) 07:29:28.96 0
オリバー・カーンのようなネアンデルタール人が、
ビンラディンのようなクロマニヨン人に滅ぼされたのが歴史の真相。
欧米人たちもそのことは認識し始めたようだ。
ビンラディンのようなクロマニヨン人に滅ぼされたのが歴史の真相。
欧米人たちもそのことは認識し始めたようだ。
643 :世界@名無史さん:2014/01/04(土) 11:03:14.76 0
ビンラディんというよりもゾマホン似か
644 :世界@名無史さん:2014/01/04(土) 11:28:01.96 0
645 :世界@名無史さん:2014/01/04(土) 12:44:50.24 0
ビンラディンも長いヒゲ剃って洋服着たら、少し日焼けした白人くらいには見えると思う
646 :世界@名無史さん:2014/01/04(土) 13:13:33.44 0
>>645
その日焼けした皮膚の色が初期コーカソイドの色(もう少し黒いかも)だろうね。
脱黒色化したのは3万年ぐらい前なのかなあ。
現在で言うとイエーメン・南部エジプトぐらいの感じ。
もちろんこれは個人的想像でしかないけどね。
その日焼けした皮膚の色が初期コーカソイドの色(もう少し黒いかも)だろうね。
脱黒色化したのは3万年ぐらい前なのかなあ。
現在で言うとイエーメン・南部エジプトぐらいの感じ。
もちろんこれは個人的想像でしかないけどね。
647 :世界@名無史さん:2014/01/04(土) 16:37:30.57 0
>>644
今のイエメン人は黒人の混血だろ
今のイエメン人は黒人の混血だろ
648 :世界@名無史さん:2014/01/04(土) 16:42:38.93 0
649 :世界@名無史さん:2014/01/05(日) 21:45:46.75 0
イエメンは北緯12度から20度だよな
それぽっち北なだけでも脱色するのか?
イエメンは今の色が限界だと思うな
それぽっち北なだけでも脱色するのか?
イエメンは今の色が限界だと思うな
650 :世界@名無史さん:2014/01/05(日) 22:14:53.88 0
平安時代でもあの絵の顔は不細工だと思うぞw
日本人の顔は1500年位前からあまり変わってないw
日本人の顔は1500年位前からあまり変わってないw
652 :世界@名無史さん:2014/01/06(月) 04:39:10.87 0
>>649
高地だけどね。
高地だけどね。
653 :世界@名無史さん:2014/01/06(月) 22:47:58.84 0
>>651 引き目鉤鼻の下膨れの典型的な日本画の人物か
ことよろ
黒い牛飼いだったと思うが出てこないな
黒い牛飼いだったと思うが出てこないな
代わりにこれで
【考古学】ウマの家畜化9000年前か、サウジアラビアで遺跡発見
http://toki.2ch.net/test/read.cgi/scienceplus/1314714359/1
1 名前:一般人φ ★[] 投稿日:2011/08/30(火) 23:25:59.99 ID:???
【8月29日 AFP】およそ9000年前に人類がウマを家畜化していたことを示す痕跡が、サウジアラビアで発見された。
定説を4000年さかのぼる新発見は24日夜、サウジアラビアの博物館・古文化局のアリ・ガッバン
(Ali al-Ghabban)副局長が会見で発表した。「これまでの研究では、人類がウマを家畜化したのは5000年前の
中央アジアだとされてきたが、今回の発見は、9000年以上前にアラビア半島(Arabian Peninsula)で最初に
家畜化されていたことを示している」
家畜化の証拠が見つかったのは、同国南西部アシール(Asir)州の州都アブハ(Abha)近くの遺跡。この地方は、
古文化学では「幸福のアラビア(Arabia Felix)」として知られる古代文明が栄えていた。ガッバン副局長によると、
この文明では「これまで知られている手法とは全く異なる死体防腐処理方法」が用いられていたという。
かつてアラビア半島が湿潤で肥よくだった頃に河床だった谷間で発見された遺跡からは、ヤギやイヌ、タカ
などの像に加え、高さ1メートルのウマの半身像があった。「これほどの寸法の動物像は、この時代としては世界の
どこでも発見されていない」とガッバン副局長。矢じりや石器、織物道具、穀類をすり潰すためのすり鉢なども
見つかっており、文明の発達ぶりが示唆されている。(c)AFP
【考古学】ウマの家畜化9000年前か、サウジアラビアで遺跡発見
http://toki.2ch.net/test/read.cgi/scienceplus/1314714359/1
1 名前:一般人φ ★[] 投稿日:2011/08/30(火) 23:25:59.99 ID:???
【8月29日 AFP】およそ9000年前に人類がウマを家畜化していたことを示す痕跡が、サウジアラビアで発見された。
定説を4000年さかのぼる新発見は24日夜、サウジアラビアの博物館・古文化局のアリ・ガッバン
(Ali al-Ghabban)副局長が会見で発表した。「これまでの研究では、人類がウマを家畜化したのは5000年前の
中央アジアだとされてきたが、今回の発見は、9000年以上前にアラビア半島(Arabian Peninsula)で最初に
家畜化されていたことを示している」
家畜化の証拠が見つかったのは、同国南西部アシール(Asir)州の州都アブハ(Abha)近くの遺跡。この地方は、
古文化学では「幸福のアラビア(Arabia Felix)」として知られる古代文明が栄えていた。ガッバン副局長によると、
この文明では「これまで知られている手法とは全く異なる死体防腐処理方法」が用いられていたという。
かつてアラビア半島が湿潤で肥よくだった頃に河床だった谷間で発見された遺跡からは、ヤギやイヌ、タカ
などの像に加え、高さ1メートルのウマの半身像があった。「これほどの寸法の動物像は、この時代としては世界の
どこでも発見されていない」とガッバン副局長。矢じりや石器、織物道具、穀類をすり潰すためのすり鉢なども
見つかっており、文明の発達ぶりが示唆されている。(c)AFP
欧州人の祖先は3系統
大昔のヨーロッパ人のゲノムから祖先がごちゃ混ぜであることが明らかになった
http://yuihaga.blog.fc2.com/blog-entry-278.html
(から抜粋)
Ewen Callaway
02 January 2014
この結論は8,000年前の狩猟採集民(ルクセンブルクからの1男性とスウェーデンからの7個体)のゲノムおよび7,500年前のドイツにいた女性のゲノムから導かれた。
テュービンゲン大学(ドイツ)のヨハネス・クラウゼ(Johannes Krause)とハーバード大学医学大学院(マサチューセッツ州ボストン)のデイヴィッド・ライヒ(David Reich)が主導した本分析は、
2013年12月23日に生物学のプレプリントウェブサイトbioRxiv.orgに投稿された。
この研究結果はまだ査読誌には発表されていない。
ミルクか穀物か
新しい研究は、現生人類で形質との繋がりが分かっているDNAの変異に基づいて、初期ヨーロッパ人の姿を描き出す。
著者たちが解読した遺伝子の配列によると、ルクセンブルクとスペインの個体は肌の色は濃かったが、青い目を持ち、狩猟採集民として知られるグループに属することが分かった。
いっぽうで、ドイツの女性は茶色い目と薄い色の肌を持ち、農耕を発達させたことが分かっている中東の人びとのグループに近縁だった。
だが、ルクセンブルクの狩猟採集民とドイツの農耕民の両方が唾液中のデンプンを分解する遺伝子のコピーを複数持っていた。
これは農耕生活に特徴的な穀物ばかりの食事への適応であると提唱されている特徴だ。
それに対して、いずれの人びともミルクの中に見られる糖、ラクトースを消化する能力を持っていなかった。
これはウシの家畜化の後に中東で出現し、後にヨーロッパに広まった特徴だ。
大昔のヨーロッパ人のゲノムから祖先がごちゃ混ぜであることが明らかになった
http://yuihaga.blog.fc2.com/blog-entry-278.html
(から抜粋)
Ewen Callaway
02 January 2014
この結論は8,000年前の狩猟採集民(ルクセンブルクからの1男性とスウェーデンからの7個体)のゲノムおよび7,500年前のドイツにいた女性のゲノムから導かれた。
テュービンゲン大学(ドイツ)のヨハネス・クラウゼ(Johannes Krause)とハーバード大学医学大学院(マサチューセッツ州ボストン)のデイヴィッド・ライヒ(David Reich)が主導した本分析は、
2013年12月23日に生物学のプレプリントウェブサイトbioRxiv.orgに投稿された。
この研究結果はまだ査読誌には発表されていない。
ミルクか穀物か
新しい研究は、現生人類で形質との繋がりが分かっているDNAの変異に基づいて、初期ヨーロッパ人の姿を描き出す。
著者たちが解読した遺伝子の配列によると、ルクセンブルクとスペインの個体は肌の色は濃かったが、青い目を持ち、狩猟採集民として知られるグループに属することが分かった。
いっぽうで、ドイツの女性は茶色い目と薄い色の肌を持ち、農耕を発達させたことが分かっている中東の人びとのグループに近縁だった。
だが、ルクセンブルクの狩猟採集民とドイツの農耕民の両方が唾液中のデンプンを分解する遺伝子のコピーを複数持っていた。
これは農耕生活に特徴的な穀物ばかりの食事への適応であると提唱されている特徴だ。
それに対して、いずれの人びともミルクの中に見られる糖、ラクトースを消化する能力を持っていなかった。
これはウシの家畜化の後に中東で出現し、後にヨーロッパに広まった特徴だ。
>>656
多様な移住
新しいデータと現在の個体の遺伝配列との比較によって、ヨーロッパ諸国の現居住民がこれら3つのグループの混成であることが分かった。
たとえば、スコットランド人とエストニア人はそれ以外のサンプルを採られたどのヨーロッパの人びとより北方ユーラシア人の祖先を持っている。
また、サルデーニャ人は他のヨーロッパ人より中東農耕民に近縁だ。
新しいヨーロッパ人の古代ゲノムはアフリカから何度も初期人類の侵襲があったこともほのめかす。
クラウゼのチームの発見によると、中東の農耕民はヨーロッパ人とアジア人のグループより早くアフリカの祖先から分岐した。
このパターンへの一つの可能性のある説明は農耕民が100,000-120,000年前にイスラエルとアラビア半島の居住地に植民した人類の子孫であるというものだ。
多くの研究者たちはこれらの遺跡がアフリカからの失敗した移住であると推定していた。
人類がアフリカを離れたのが100,000年以内であることを他の証拠が示していたためだ。
多様な移住
新しいデータと現在の個体の遺伝配列との比較によって、ヨーロッパ諸国の現居住民がこれら3つのグループの混成であることが分かった。
たとえば、スコットランド人とエストニア人はそれ以外のサンプルを採られたどのヨーロッパの人びとより北方ユーラシア人の祖先を持っている。
また、サルデーニャ人は他のヨーロッパ人より中東農耕民に近縁だ。
新しいヨーロッパ人の古代ゲノムはアフリカから何度も初期人類の侵襲があったこともほのめかす。
クラウゼのチームの発見によると、中東の農耕民はヨーロッパ人とアジア人のグループより早くアフリカの祖先から分岐した。
このパターンへの一つの可能性のある説明は農耕民が100,000-120,000年前にイスラエルとアラビア半島の居住地に植民した人類の子孫であるというものだ。
多くの研究者たちはこれらの遺跡がアフリカからの失敗した移住であると推定していた。
人類がアフリカを離れたのが100,000年以内であることを他の証拠が示していたためだ。
これはキタな
ちなみにひたすら貼ってるのは俺だけなんだが
アラビア含めても問題ないよね?
ちなみにひたすら貼ってるのは俺だけなんだが
アラビア含めても問題ないよね?
>>657
このあたり予定調和
現生人類の出アフリカの時期の見直し
http://sicambre.at.webry.info/201101/article_29.html
(抜粋)
この研究では、アラブ首長国連邦のジェベル=ファーヤ遺跡で発見された石器が報告されています。
その石器の年代は10万年以上前であり、要約を読むかぎり、人骨は共伴していないようですが、アフリカ北東部の中期石器時代の石器との類似性が指摘されています。
このことから、10万年以上前ジェベル=ファーヤ遺跡の石器文化の担い手は解剖学的現代人であり、
後期石器時代や上部旧石器時代移行のような技術革新がなくとも、現生人類はアフリカからアラビア半島へと進出できたのではないか、と推測されています。
ただ、現在に残る石器には現れない何らかの社会的革新が原動力となった可能性も考慮に入れるべきではないだろうか、と私は考えています。
ジェベル=ファーヤ遺跡の石器の年代は、海洋酸素同位体ステージ6〜5の移行期にかけてのことであり、
寒冷なステージ6において海面が低下したことも、現生人類のアフリカからアラビア半島への進出を容易にしたのではないか、とも指摘されています。
当時のアラビア半島は現在よりも降水量がかなり多く、多数の植物に覆われ、湖と川が網状に広がっていたと推測され、
アラビア半島での現生人類の生活は比較的容易であり、現生人類がここからさらに西アジアや南アジアへと進出していったのではないか、と推測されています。
このあたり予定調和
現生人類の出アフリカの時期の見直し
http://sicambre.at.webry.info/201101/article_29.html
(抜粋)
この研究では、アラブ首長国連邦のジェベル=ファーヤ遺跡で発見された石器が報告されています。
その石器の年代は10万年以上前であり、要約を読むかぎり、人骨は共伴していないようですが、アフリカ北東部の中期石器時代の石器との類似性が指摘されています。
このことから、10万年以上前ジェベル=ファーヤ遺跡の石器文化の担い手は解剖学的現代人であり、
後期石器時代や上部旧石器時代移行のような技術革新がなくとも、現生人類はアフリカからアラビア半島へと進出できたのではないか、と推測されています。
ただ、現在に残る石器には現れない何らかの社会的革新が原動力となった可能性も考慮に入れるべきではないだろうか、と私は考えています。
ジェベル=ファーヤ遺跡の石器の年代は、海洋酸素同位体ステージ6〜5の移行期にかけてのことであり、
寒冷なステージ6において海面が低下したことも、現生人類のアフリカからアラビア半島への進出を容易にしたのではないか、とも指摘されています。
当時のアラビア半島は現在よりも降水量がかなり多く、多数の植物に覆われ、湖と川が網状に広がっていたと推測され、
アラビア半島での現生人類の生活は比較的容易であり、現生人類がここからさらに西アジアや南アジアへと進出していったのではないか、と推測されています。
>>599
・MA-1 Y 24,000y
・AG-2 X 17,000y
>>656
“Stuttgart”, a ~7,500 year old individual
found in Stuttgart in southern Germany who was buried in the context of artifacts from the first
widespread Neolithic farming culture of central and northern Europe, the Linearbandkeramik (LBK).
“Loschbour”, an ~8,000 year old individual
found in the Loschbour rock shelter in Heffingen Luxembourg, from a skeleton that was
discovered in the context of Mesolithic hunter-gatherer artifacts (SI1; SI2).
We also sequenced DNA from seven ~8,000 year old remains from Mesolithic hunter-gatherers from
the Motala site in southern Sweden, with the highest coverage individual (Motala12) at 2.4-fold
Stuttgart belonged to
mtDNA haplogroup T2, typical of Neolithic Europeans
, while Loschbour and all Motala
individuals belonged to haplogroups U5 and U2, typical of pre-agricultural Europeans (SI4).
Based on the ratio of sequences aligning to chromosomes X and Y, we infer that
Stuttgart was female
while Loschbour and five Motala individuals were male (SI5).
Loschbour and four Motala males belonged to Y-chromosome haplogroup I,
showing that this was a predominant
haplogroup in pre-agricultural northern Europeans (SI5).
・MA-1 Y 24,000y
・AG-2 X 17,000y
>>656
“Stuttgart”, a ~7,500 year old individual
found in Stuttgart in southern Germany who was buried in the context of artifacts from the first
widespread Neolithic farming culture of central and northern Europe, the Linearbandkeramik (LBK).
“Loschbour”, an ~8,000 year old individual
found in the Loschbour rock shelter in Heffingen Luxembourg, from a skeleton that was
discovered in the context of Mesolithic hunter-gatherer artifacts (SI1; SI2).
We also sequenced DNA from seven ~8,000 year old remains from Mesolithic hunter-gatherers from
the Motala site in southern Sweden, with the highest coverage individual (Motala12) at 2.4-fold
Stuttgart belonged to
mtDNA haplogroup T2, typical of Neolithic Europeans
, while Loschbour and all Motala
individuals belonged to haplogroups U5 and U2, typical of pre-agricultural Europeans (SI4).
Based on the ratio of sequences aligning to chromosomes X and Y, we infer that
Stuttgart was female
while Loschbour and five Motala individuals were male (SI5).
Loschbour and four Motala males belonged to Y-chromosome haplogroup I,
showing that this was a predominant
haplogroup in pre-agricultural northern Europeans (SI5).
661 :世界@名無史さん:2014/01/08(水) 23:03:42.03 0
>>657 サルデーニャ人はY-DNAでI系統が多く、旧石器時代の人類の直系の子孫と言うのが直近の説だったけど、また変わったのか
あるいは10万年以上前に中東に移住し、そこから農耕を開始する前の更新世のうちにヨーロッパに渡ったと考えれば矛盾はないのか
スコットランド人とエストニア人が多く持つ北方ユーラシア人の祖先って、ひょっとしてY-DNAのN系統とほぼ同義?
スコットランドに多いとは聞かないけど、海の妖精セルキーが、アザラシの姿から人に化けて人と交わって子を成した、と言う伝説も何かの示唆かな
一番近くのモンゴロイド・エスキモーにはN系統は少ないかほとんどないようだけど
あるいは10万年以上前に中東に移住し、そこから農耕を開始する前の更新世のうちにヨーロッパに渡ったと考えれば矛盾はないのか
スコットランド人とエストニア人が多く持つ北方ユーラシア人の祖先って、ひょっとしてY-DNAのN系統とほぼ同義?
スコットランドに多いとは聞かないけど、海の妖精セルキーが、アザラシの姿から人に化けて人と交わって子を成した、と言う伝説も何かの示唆かな
一番近くのモンゴロイド・エスキモーにはN系統は少ないかほとんどないようだけど
>>661
その北方ユーラシア人の祖先って、MA-1とAG-2 だよ。
その北方ユーラシア人の祖先って、MA-1とAG-2 だよ。
666 :世界@名無史さん:2014/01/08(水) 23:51:57.66 0
あの、イエメンに限らないけどアラブ人は黒人奴隷を随分とアフリカから連れて来てて
混血まるけなんだけど。だからイエメンだかにネグリトっぽい集団もいるらしいけど
古代にアフリカを出たというより近代にアフリカから来たものと区別がつかないとか
でいうか、7万年前に出てから全然出て無いわけないだろ。ナイル川を北上ものできるし
混血まるけなんだけど。だからイエメンだかにネグリトっぽい集団もいるらしいけど
古代にアフリカを出たというより近代にアフリカから来たものと区別がつかないとか
でいうか、7万年前に出てから全然出て無いわけないだろ。ナイル川を北上ものできるし
668 :世界@名無史さん:2014/01/09(木) 00:02:43.73 0
アラブ人には黒人に極めて違いのと白人に極めて近いのがいる。
キューバやプエルトリコの黒人色の強いヒスパニックと同じような感じ。
キューバやプエルトリコの黒人色の強いヒスパニックと同じような感じ。
謎の隕石を追う 砂漠が燃えた日
http://www.nikkei-science.net/modules/flash/index.php?id=199902_020
日経サイエンス 1999年2月号
J. C. ウィン
E. M. シューメーカー(ともに米国地質調査所)
アラビア半島の地図を見ると,内陸部に町も村もない広大な空白地帯が存在することがわかる。
サウジアラビアのルブ・アル・ハリ砂漠だ。その砂の海の奥深くに非常に奇妙な場所がある。
広さ0.5km2ほどの地域で,周囲は砂ばかりなのに,そこだけ黒っぽいガラスや白い岩石,鉄の破片などが散らばっている。
この不思議な地域の話は1932年,英国の探検家,ジョン=アブドゥラ=フィルビーによって初めて欧米世界に伝えられた。
もっとも,アラビア半島に古くから住む放浪の民ベドウィン族の間では,この地は「鉄のある場所(Theiron things)」として何世代にもわたって語り継がれ,
いにしえの王国の都ウバールの跡とも言い伝えられていた。
彼は旅行記の中で,この土地の名前を「ウバール」ではなく,「ワバール」と聞き書きで記した。
以来,この地は一般に「ワバール」という名前で通るようになった。しかし,フィルビーが発見したのは失われたウバールの町ではなかった。
ただ,確かにそこは空からの災厄に見舞われた跡だった。隕石が衝突したのだ。
ワバールは隕石探査における特別な場所となっている。これまで地球に衝突した隕石のほとんどは,表面が土や岩石で覆われた固い大陸表面か,さもなければ海などに落ちている。
これに対して,ワバールでは,落ちたところが世界でも指折りの砂の海だった点が他と大きく異なる。
この乾燥した別世界は,地質学的に見て,隕石衝突の跡が,地上で最もよく保存された場所といえるだろう。
ではいったいワバールでどんなことが実際に起きたのか,私たちは3度に及ぶルブ・アル・ハリ砂漠への大変な調査旅行によって,衝突当時の模様を再現することに成功した。(本文から)
http://www.nikkei-science.net/modules/flash/index.php?id=199902_020
日経サイエンス 1999年2月号
J. C. ウィン
E. M. シューメーカー(ともに米国地質調査所)
アラビア半島の地図を見ると,内陸部に町も村もない広大な空白地帯が存在することがわかる。
サウジアラビアのルブ・アル・ハリ砂漠だ。その砂の海の奥深くに非常に奇妙な場所がある。
広さ0.5km2ほどの地域で,周囲は砂ばかりなのに,そこだけ黒っぽいガラスや白い岩石,鉄の破片などが散らばっている。
この不思議な地域の話は1932年,英国の探検家,ジョン=アブドゥラ=フィルビーによって初めて欧米世界に伝えられた。
もっとも,アラビア半島に古くから住む放浪の民ベドウィン族の間では,この地は「鉄のある場所(Theiron things)」として何世代にもわたって語り継がれ,
いにしえの王国の都ウバールの跡とも言い伝えられていた。
彼は旅行記の中で,この土地の名前を「ウバール」ではなく,「ワバール」と聞き書きで記した。
以来,この地は一般に「ワバール」という名前で通るようになった。しかし,フィルビーが発見したのは失われたウバールの町ではなかった。
ただ,確かにそこは空からの災厄に見舞われた跡だった。隕石が衝突したのだ。
ワバールは隕石探査における特別な場所となっている。これまで地球に衝突した隕石のほとんどは,表面が土や岩石で覆われた固い大陸表面か,さもなければ海などに落ちている。
これに対して,ワバールでは,落ちたところが世界でも指折りの砂の海だった点が他と大きく異なる。
この乾燥した別世界は,地質学的に見て,隕石衝突の跡が,地上で最もよく保存された場所といえるだろう。
ではいったいワバールでどんなことが実際に起きたのか,私たちは3度に及ぶルブ・アル・ハリ砂漠への大変な調査旅行によって,衝突当時の模様を再現することに成功した。(本文から)
670 :世界@名無史さん:2014/01/09(木) 11:21:18.73 0
>>666 オッペンハイマーは10万年前にパレスチナに出た集団は死滅したと考えてるけど、ナイル河谷にいたら環境の激変を乗り越えられた気もする
パレスチナにまで行ったんだから、途中のエジプトにとどまった集団がいたとしてもおかしくない
パレスチナにまで行ったんだから、途中のエジプトにとどまった集団がいたとしてもおかしくない
671 :世界@名無史さん:2014/01/09(木) 13:06:25.67 0
ペルシャ湾も緑豊かな陸地だったわけで7万年前から氷河期が終わる一万年前
までコーカソイドの先祖やドラビタ人の先祖達が住んでいただろう。
ヨーロッパに4万年前に入った人類はアボリジニに頭骨は似てるから黒人らしい
コーカソイドはヨーロッパじゃなくペルシャ湾を含む中東で進化したんじゃないか?
http://docs.docstoc.com/pdf/5428307/3bd3298f-7199-48be-8522-854d1871a992.pdf
までコーカソイドの先祖やドラビタ人の先祖達が住んでいただろう。
ヨーロッパに4万年前に入った人類はアボリジニに頭骨は似てるから黒人らしい
コーカソイドはヨーロッパじゃなくペルシャ湾を含む中東で進化したんじゃないか?
http://docs.docstoc.com/pdf/5428307/3bd3298f-7199-48be-8522-854d1871a992.pdf
672 :世界@名無史さん:2014/01/09(木) 19:20:59.27 0
白色化の前に高鼻化が起きたか。縮毛から波状毛の変化もここで起きたのかな?あるいはアボリジニとかタミル人から類推すると、波状毛が人類の本来の姿?
でも世界一古いアンダマン先住民やコイサンは縮毛だな
でも世界一古いアンダマン先住民やコイサンは縮毛だな
通説はアナトリアからの移住だったと思うんだけど
これはこれで異様におもしろいなww
これはこれで異様におもしろいなww
欧米人が世界を征服したからそれが美の基準になっただけ
また肌の白さも顔の彫りの深さも足の長さも全部生存には不利な特徴でしかないので
一部地域でしか多数派を形成し得ず結果的に希少価値と見做されるようになっているのだろう
また肌の白さも顔の彫りの深さも足の長さも全部生存には不利な特徴でしかないので
一部地域でしか多数派を形成し得ず結果的に希少価値と見做されるようになっているのだろう
676 :世界@名無史さん:2014/01/09(木) 23:08:28.92 0
モンゴロイドが世界を征服してもこれが美の基準にならんだろ
http://blogimg.goo.ne.jp/user_image/05/04/d6ce1c3f56e107bea2c03723f991436a.jpg
http://blogimg.goo.ne.jp/user_image/05/04/d6ce1c3f56e107bea2c03723f991436a.jpg
{kind=link}
今は年を取っているからなw 若い頃は…まあw
でも、ルイスフロイスも戦国時代の日本人の美の価値観で「目が細い方が美しいとされる」と明記しているしなあ。
でも、ルイスフロイスも戦国時代の日本人の美の価値観で「目が細い方が美しいとされる」と明記しているしなあ。
678 :世界@名無史さん:2014/01/09(木) 23:30:32.34 0
アフリカの黒人やアンダマンの黒人ってなんで短髪なん?
火さえ知らなかったアンダマンやブッシュマンが床屋に行ってるとは思えないし
チンパンジーは短髪だから逆に長髪に成った出アフリカ人が長髪なのが不思議
これはコーカソイドもモンゴロイドもアイヌなど古代人も同じ
火さえ知らなかったアンダマンやブッシュマンが床屋に行ってるとは思えないし
チンパンジーは短髪だから逆に長髪に成った出アフリカ人が長髪なのが不思議
これはコーカソイドもモンゴロイドもアイヌなど古代人も同じ
679 :世界@名無史さん:2014/01/09(木) 23:43:31.21 0
【朝日新聞】 「国家は初中等学校における歴史教育を廃止すべきだ…歴史認識は、国家間だけでなく専門家の間にも対立がある」★3
http://uni.2ch.net/test/read.cgi/newsplus/1389262131/
【政治】 「東京都知事になったら、ソウル・北京と平和自治体会議したい」 前回の都知事選で100万票近く獲得の宇都宮健児氏★2
http://uni.2ch.net/test/read.cgi/newsplus/1389234961/
【社会】「レイシスト(人種差別)しばき隊」関係者が「友人」と明かす 身柄確保の杉本容疑者は何者なのか
http://uni.2ch.net/test/read.cgi/newsplus/1389269501/
http://uni.2ch.net/test/read.cgi/newsplus/1389262131/
【政治】 「東京都知事になったら、ソウル・北京と平和自治体会議したい」 前回の都知事選で100万票近く獲得の宇都宮健児氏★2
http://uni.2ch.net/test/read.cgi/newsplus/1389234961/
【社会】「レイシスト(人種差別)しばき隊」関係者が「友人」と明かす 身柄確保の杉本容疑者は何者なのか
http://uni.2ch.net/test/read.cgi/newsplus/1389269501/
680 :世界@名無史さん:2014/01/10(金) 01:51:22.15 0
どの分野にも行っても嫌韓ネタかあるわけで、もうスレ違とかのレベルじゃない。
これは日本人全体の意識が変わりつつある兆候で、韓国が叩けば叩くほど嫌韓が増える。
韓国人は一体何をしたいのか? アメリカでどんだけ黒人に嫌われようがお構いなしだから
特に日本に対しては、嫌われようが何だろうがイケイケなんだろうな。
これは日本人全体の意識が変わりつつある兆候で、韓国が叩けば叩くほど嫌韓が増える。
韓国人は一体何をしたいのか? アメリカでどんだけ黒人に嫌われようがお構いなしだから
特に日本に対しては、嫌われようが何だろうがイケイケなんだろうな。
コピペしているだけだろw >>680もコピペ?
>>682
島の名前は勘違いだったので
合ってるんだろうな
http://en.wikipedia.org/wiki/Guanches
Origins
Genetic evidence shows that northern African peoples (possibly descendants of the Capsian culture) made a significant contribution to the aboriginal population of the Canaries following desertification of the Sahara at some point after 6000 BCE.
Linguistic evidence suggests ties between the Guanche language and the Berber languages of North Africa,
particularly when comparing numeral systems.[6][8]
Research into the genetics of the Guanche population have led to the conclusion that they share an ancestry with Berber peoples.[9]
The islands were visited by a number of peoples within recorded history.
The Numidians, Phoenicians, and Carthaginians knew of the islands and made frequent visits,[10] including expeditions dispatched from Mogador by Juba.[11]
The Romans occupied northern Africa and visited the Canaries between the 1st and 4th centuries CE,
judging from Roman artifacts found on the island of Lanzarote.
These show that Romans did trade with the Canaries, though there is no evidence of their ever settling there.[12]
Archaeology of the Canaries seem to reflect diverse levels of technology, some differing from the Neolithic culture that was encountered at the time of conquest.
It is thought that the arrival of the aborigines to the archipelago led to the extinction of some big reptiles and insular mammals,
for example, the giant lizard Gallotia goliath (which managed to reach up to a meter in length) and Canariomys bravoi, the giant rat of Tenerife.
Population genetics にはハプログループもいろいろと載ってる
島の名前は勘違いだったので
合ってるんだろうな
http://en.wikipedia.org/wiki/Guanches
Origins
Genetic evidence shows that northern African peoples (possibly descendants of the Capsian culture) made a significant contribution to the aboriginal population of the Canaries following desertification of the Sahara at some point after 6000 BCE.
Linguistic evidence suggests ties between the Guanche language and the Berber languages of North Africa,
particularly when comparing numeral systems.[6][8]
Research into the genetics of the Guanche population have led to the conclusion that they share an ancestry with Berber peoples.[9]
The islands were visited by a number of peoples within recorded history.
The Numidians, Phoenicians, and Carthaginians knew of the islands and made frequent visits,[10] including expeditions dispatched from Mogador by Juba.[11]
The Romans occupied northern Africa and visited the Canaries between the 1st and 4th centuries CE,
judging from Roman artifacts found on the island of Lanzarote.
These show that Romans did trade with the Canaries, though there is no evidence of their ever settling there.[12]
Archaeology of the Canaries seem to reflect diverse levels of technology, some differing from the Neolithic culture that was encountered at the time of conquest.
It is thought that the arrival of the aborigines to the archipelago led to the extinction of some big reptiles and insular mammals,
for example, the giant lizard Gallotia goliath (which managed to reach up to a meter in length) and Canariomys bravoi, the giant rat of Tenerife.
Population genetics にはハプログループもいろいろと載ってる
686 :世界@名無史さん:2014/01/10(金) 04:18:30.68 0
性淘汰
ヨーロッパというか中東にはエデンの園があって(ペルシャ湾とも言われてる)
有力者は美人を集めてハーレムを作った。豊かなエデンの園でのハーレムでは
美人美男がどんどん増えたがペルシャ湾は氷河期の終焉で大洪水、海となった。
一方、3万年ぐらい前から寒冷地帯に入った東ユーラシアの人類は全身を毛皮を
まとい顔姿は生きて行く上で二の次になり、それで性淘汰も起きず寒冷地に適応
した平たい顔つきのホモサピエンスで最も異様な顔付きのモンゴロイドとなった。
一方、ヨーロッパの北欧の人類は氷河に覆われていたので中東や中央アジアから
入ったのは氷河期が終わったせいぜい1万年前後なので寒冷適応する時間もなかった。
異論は認める。
ヨーロッパというか中東にはエデンの園があって(ペルシャ湾とも言われてる)
有力者は美人を集めてハーレムを作った。豊かなエデンの園でのハーレムでは
美人美男がどんどん増えたがペルシャ湾は氷河期の終焉で大洪水、海となった。
一方、3万年ぐらい前から寒冷地帯に入った東ユーラシアの人類は全身を毛皮を
まとい顔姿は生きて行く上で二の次になり、それで性淘汰も起きず寒冷地に適応
した平たい顔つきのホモサピエンスで最も異様な顔付きのモンゴロイドとなった。
一方、ヨーロッパの北欧の人類は氷河に覆われていたので中東や中央アジアから
入ったのは氷河期が終わったせいぜい1万年前後なので寒冷適応する時間もなかった。
異論は認める。
687 :世界@名無史さん:2014/01/10(金) 04:51:08.05 O
シュメール人はコーカソイドだが遺伝子は何だろうね
シュメール文明は5000年前に突然現れたかんじで
すでに文字もあり文学もあった。それが旧約聖書によく似てて
エデンの園も大洪水もノアの箱舟も出てくる
あれはペルシャ湾の終焉が口授されたとも言われる
シュメール文明は5000年前に突然現れたかんじで
すでに文字もあり文学もあった。それが旧約聖書によく似てて
エデンの園も大洪水もノアの箱舟も出てくる
あれはペルシャ湾の終焉が口授されたとも言われる
688 :世界@名無史さん:2014/01/10(金) 22:50:14.73 0
>>683 グイマー(魔境グンマーではない)のピラミッドは誰が作ったんだろうな。スペイン人と接触したグアンチェ人が石器時代の暮らしだったことを考えると、
彼らではなさそう。
彼らではなさそう。
689 :世界@名無史さん:2014/01/10(金) 22:57:57.94 0
群馬辺りは高句麗人が移住して子孫が多いらしい。
埼玉も高麗川とかの地名があるから高句麗人の子孫が多いだろうな。
埼玉も高麗川とかの地名があるから高句麗人の子孫が多いだろうな。
俺も、記憶と記録、時代と自大、が錯綜思想になるんで
気になったことをメモっておくけど
・ハンノの航海のとき島は無人だった?
・世界ふしぎ発見で取り上げられていたはずだが記録がない?
http://www.eorc.jaxa.jp/imgdata/topics/2010/tp101129.html
http://en.wikipedia.org/wiki/Museo_de_la_Naturaleza_y_el_Hombre
Featured Pieces[edit]
Zanata Stone.
Mummy of San Andres.The museum features a formidable collection of mummies, ornaments, models of aboriginal settlements and pottery.
The history ocvered ranges from the beginnings of the Guanche culture to the Conquest of the Canary Islands. Some of its key parts are:
Guanche mummies. Among which the famous Mummy of San Andres
Zanata Stone (rock with Berber-Punic inscriptions)
Guanche ceramics and personal items
Mediterranean and Aegean amphoras found on the shores of the Canary Islands
Thirteen Egyptian ceramics dated to 5700 years old, one of which is the oldest piece of this civilization in a Spanish museum.[7]
ミイラはある。
エジプトと地中海とエーゲ海との交流はあった。
化石人骨は記述がない。
>>683
mtDNA haplogroup U subclade U6b1 is Canarian-specific[13] and is the most common mtDNA haplogroup found in aboriginal Guanche archaeological burial sites.[9
>>688
ヘイエルダールらしいww
気になったことをメモっておくけど
・ハンノの航海のとき島は無人だった?
・世界ふしぎ発見で取り上げられていたはずだが記録がない?
http://www.eorc.jaxa.jp/imgdata/topics/2010/tp101129.html
http://en.wikipedia.org/wiki/Museo_de_la_Naturaleza_y_el_Hombre
Featured Pieces[edit]
Zanata Stone.
Mummy of San Andres.The museum features a formidable collection of mummies, ornaments, models of aboriginal settlements and pottery.
The history ocvered ranges from the beginnings of the Guanche culture to the Conquest of the Canary Islands. Some of its key parts are:
Guanche mummies. Among which the famous Mummy of San Andres
Zanata Stone (rock with Berber-Punic inscriptions)
Guanche ceramics and personal items
Mediterranean and Aegean amphoras found on the shores of the Canary Islands
Thirteen Egyptian ceramics dated to 5700 years old, one of which is the oldest piece of this civilization in a Spanish museum.[7]
ミイラはある。
エジプトと地中海とエーゲ海との交流はあった。
化石人骨は記述がない。
>>683
mtDNA haplogroup U subclade U6b1 is Canarian-specific[13] and is the most common mtDNA haplogroup found in aboriginal Guanche archaeological burial sites.[9
>>688
ヘイエルダールらしいww
>>656
一つのポイントはスカンジナビアの古代先住民が
現代の住民とあんまり似てないとこか
ギリシャからは
ギリシャ→トスカーナ→ベルガモ の流れと
ギリシャ→ブルガリア、アルバニア→クロアチア の流れ
2つに分かれて見えるんだが
でもギリシャ自体は一つの分布なんだよな
アカイア、ドーリア、アイオリスとかに別れるかと思ったのに
一つのポイントはスカンジナビアの古代先住民が
現代の住民とあんまり似てないとこか
ギリシャからは
ギリシャ→トスカーナ→ベルガモ の流れと
ギリシャ→ブルガリア、アルバニア→クロアチア の流れ
2つに分かれて見えるんだが
でもギリシャ自体は一つの分布なんだよな
アカイア、ドーリア、アイオリスとかに別れるかと思ったのに
>>660
Table S4.1. Haplogroup
Sample haplogroup
Loschbour U5b1a
Motala 1 U5a1
Motala 2 U2e1
Motala 3 U5a1
Motala 4 U5a2d
Motala 6 U5a2d
Motala 9 U5a2
Motala 12 U2e1
Stuttgart T2c1d1
Table S4.1. Haplogroup
Sample haplogroup
Loschbour U5b1a
Motala 1 U5a1
Motala 2 U2e1
Motala 3 U5a1
Motala 4 U5a2d
Motala 6 U5a2d
Motala 9 U5a2
Motala 12 U2e1
Stuttgart T2c1d1
>>660
Table S2.3: Number of autosomal SNPs with an allele call
Sample SNPs†
Motala12 411,453
Motala_merge* 352,966
Skoglund_farmer 4,548
Skoglund_mergeHG 18,261
Tyrolean Iceman 518,229
La Brana 1 9,868
La Brana 2 4,525
* Motala_merge includes reads from five Motala samples (all except Motala3 and Motala12).
(EEF) Early European Farmers
Skoglund_farmer
Tyrolean Iceman
Stuttgart
(WHG) West European Hunter-Gatherers
Loschbour
LaBrana1
LaBrana2
(ANE) Ancient North Eurasians
MA1
AG2
Table S2.3: Number of autosomal SNPs with an allele call
Sample SNPs†
Motala12 411,453
Motala_merge* 352,966
Skoglund_farmer 4,548
Skoglund_mergeHG 18,261
Tyrolean Iceman 518,229
La Brana 1 9,868
La Brana 2 4,525
* Motala_merge includes reads from five Motala samples (all except Motala3 and Motala12).
(EEF) Early European Farmers
Skoglund_farmer
Tyrolean Iceman
Stuttgart
(WHG) West European Hunter-Gatherers
Loschbour
LaBrana1
LaBrana2
(ANE) Ancient North Eurasians
MA1
AG2
>>693>>684
http://dienekes.blogspot.jp/2012/06/mesolithic-iberians-la-brana-arintero.html
A team of scientists, led by researcher Carles Lalueza-Fox from CSIC (Spanish National Research Council),
has recovered - for the first time in history - part of the genome of two individuals living in the Mesolithic Period, 7000 years ago.
Remains have been found at La Brana-Arintero site, located at Valdelugueros (Leon), Spain.
The study results, published in the Current Biology magazine, indicate that current Iberian populations don't come from these groups genetically.
http://es.wikipedia.org/wiki/Valdelugueros
http://es.wikipedia.org/wiki/La_Bra%C3%B1a
http://dienekes.blogspot.jp/2012/06/mesolithic-iberians-la-brana-arintero.html
A team of scientists, led by researcher Carles Lalueza-Fox from CSIC (Spanish National Research Council),
has recovered - for the first time in history - part of the genome of two individuals living in the Mesolithic Period, 7000 years ago.
Remains have been found at La Brana-Arintero site, located at Valdelugueros (Leon), Spain.
The study results, published in the Current Biology magazine, indicate that current Iberian populations don't come from these groups genetically.
http://es.wikipedia.org/wiki/Valdelugueros
http://es.wikipedia.org/wiki/La_Bra%C3%B1a
>>694
http://forwhattheywereweare.blogspot.jp/2012/06/more-north-iberian-epipaleolithic-mtdna.html
In this case two individuals (right) from La Brana-Arintero site in Valdelugueros (Leon province near the Asturian border),
dated to 7000 years ago, borderline with Neolithic arrival but, according to the paper, still "Mesolithic" (i.e. Epipaleolithic).
Federico Sanchez Quinto et al., Genomic Affinities of Two 7,000-Year-Old Iberian Hunter-Gatherers. Current Biology 2012. Pay per view.
The two individuals carried haplogroup U5b2c1 (I guess that they were relatives, maybe siblings). The mtDNA sequence was retrieved in full.
It is worth mentioning that there have been other recent studies of Northern Iberian pre-Neolithic mtDNA:
?M. Lacan found mtDNA H among Epipaleolithic Basques in 2011.
?M. Hervella found mtDNA H and U5 among pre-Neolithic Cantabrians and Basques respectively a couple of months ago.
http://1.bp.blogspot.com/-nJV5OmLbYgc/T-yxfcU8BjI/AAAAAAAABBc/ZJmzwwRqr8g/s1600/North+Iberia+pre-Neolithic+mtDNA+2.png
7,000BP Iberian hunter gatherer La Brana-1 had blue eyes
http://www.theapricity.com/forum/showthread.php?105677-7-000BP-Iberian-hunter-gatherer-La-Brana-1-had-blue-eyes
http://forwhattheywereweare.blogspot.jp/2012/06/more-north-iberian-epipaleolithic-mtdna.html
In this case two individuals (right) from La Brana-Arintero site in Valdelugueros (Leon province near the Asturian border),
dated to 7000 years ago, borderline with Neolithic arrival but, according to the paper, still "Mesolithic" (i.e. Epipaleolithic).
Federico Sanchez Quinto et al., Genomic Affinities of Two 7,000-Year-Old Iberian Hunter-Gatherers. Current Biology 2012. Pay per view.
The two individuals carried haplogroup U5b2c1 (I guess that they were relatives, maybe siblings). The mtDNA sequence was retrieved in full.
It is worth mentioning that there have been other recent studies of Northern Iberian pre-Neolithic mtDNA:
?M. Lacan found mtDNA H among Epipaleolithic Basques in 2011.
?M. Hervella found mtDNA H and U5 among pre-Neolithic Cantabrians and Basques respectively a couple of months ago.
http://1.bp.blogspot.com/-nJV5OmLbYgc/T-yxfcU8BjI/AAAAAAAABBc/ZJmzwwRqr8g/s1600/North+Iberia+pre-Neolithic+mtDNA+2.png
{kind=link}
7,000BP Iberian hunter gatherer La Brana-1 had blue eyes
http://www.theapricity.com/forum/showthread.php?105677-7-000BP-Iberian-hunter-gatherer-La-Brana-1-had-blue-eyes
ところで誰かバスク地方で発見された
石器時代の100体クラス人骨が見つかった遺跡の名前しらない?
石器時代の100体クラス人骨が見つかった遺跡の名前しらない?
>>693
Ancient DNA from Neolithic Sweden (Skoglund et al. 2012)
http://dienekes.blogspot.jp/2012/04/ancient-dna-from-neolithic-sweden.html
長い、、、
The Neolithic farmer sample ('Gok4')
was excavated from a megalithic burial structure in Gokhem parish,
Sweden, and has been directly 14C-dated to 4,921 ± 50 calibrated years BP (calBP), similar to the age (5,100-4,900 calBP) of the majority of other finds in the area (15).
There were no indications from the burial context suggesting that Gok4 was different from other TRB individuals (15, 16),
and strontium isotope analyses indicate that Gok4 was born less than 100 km from the megalithic structure, similar to all other analyzed TRB individuals from the area (17).
The three Neolithic hunter-gatherer samples were excavated from burial grounds with single inhumation graves on the island of Gotland, Sweden,
for which associated remains have been dated to 5300-4400 calBP (16).
http://3.bp.blogspot.com/-Ncn8vTOFEaE/T5WslfRh-bI/AAAAAAAAEyg/S5T4Oa5lFuI/s1600/fg5_online.gif
Ancient DNA from Neolithic Sweden (Skoglund et al. 2012)
http://dienekes.blogspot.jp/2012/04/ancient-dna-from-neolithic-sweden.html
長い、、、
The Neolithic farmer sample ('Gok4')
was excavated from a megalithic burial structure in Gokhem parish,
Sweden, and has been directly 14C-dated to 4,921 ± 50 calibrated years BP (calBP), similar to the age (5,100-4,900 calBP) of the majority of other finds in the area (15).
There were no indications from the burial context suggesting that Gok4 was different from other TRB individuals (15, 16),
and strontium isotope analyses indicate that Gok4 was born less than 100 km from the megalithic structure, similar to all other analyzed TRB individuals from the area (17).
The three Neolithic hunter-gatherer samples were excavated from burial grounds with single inhumation graves on the island of Gotland, Sweden,
for which associated remains have been dated to 5300-4400 calBP (16).
http://3.bp.blogspot.com/-Ncn8vTOFEaE/T5WslfRh-bI/AAAAAAAAEyg/S5T4Oa5lFuI/s1600/fg5_online.gif
{kind=link}
>>687
シュメール人はひとまず置いておくとして
サハラ平原に住んでいた人類はどんな種類の人だったと思う?
>>670
オッペンハイマーだから、そんな迂闊な言い方はしてないと思うな。
俺が読んだ人類拡散本の中で、時の試練に耐えてるのはこれぐらい。
シュメール人はひとまず置いておくとして
サハラ平原に住んでいた人類はどんな種類の人だったと思う?
>>670
オッペンハイマーだから、そんな迂闊な言い方はしてないと思うな。
俺が読んだ人類拡散本の中で、時の試練に耐えてるのはこれぐらい。
>>574
約5000年前のスカンジナビアには
南方由来の初期農耕民と、西方由来の狩猟採集民が
同時期に存在していたが、
現在のスカンジナビア人は両方とも異なっており
ブルガリア→クロアチア→ハンガリー付近を通って到達した民族に由来する。
約5000年前のスカンジナビアには
南方由来の初期農耕民と、西方由来の狩猟採集民が
同時期に存在していたが、
現在のスカンジナビア人は両方とも異なっており
ブルガリア→クロアチア→ハンガリー付近を通って到達した民族に由来する。
>>559>>608
>>607>>671
よく読んでなかったけど、こういうことらしい
http://dienekes.blogspot.jp/2014/01/slc24a5-light-skin-pigmentation-allele.html
Molecular Phylogeography of a Human Autosomal Skin Color Locus Under Natural Selection
Virtually all chromosomes carrying the A111T allele share a single 78-kb haplotype that we call C11, indicating that all instances of this mutation in human populations share a common origin.
。。。
The distributions of C11 and its parental haplotypes make it most likely that these two last steps occurred between the Middle East and the Indian subcontinent,
with the A111T mutation occurring after the split between the ancestors of Europeans and East Asians.
>>607>>671
よく読んでなかったけど、こういうことらしい
http://dienekes.blogspot.jp/2014/01/slc24a5-light-skin-pigmentation-allele.html
Molecular Phylogeography of a Human Autosomal Skin Color Locus Under Natural Selection
Virtually all chromosomes carrying the A111T allele share a single 78-kb haplotype that we call C11, indicating that all instances of this mutation in human populations share a common origin.
。。。
The distributions of C11 and its parental haplotypes make it most likely that these two last steps occurred between the Middle East and the Indian subcontinent,
with the A111T mutation occurring after the split between the ancestors of Europeans and East Asians.
>>662
ジブラルタルコースもあり得るようだww
http://en.wikipedia.org/wiki/Berber_people
Origins[edit]
Further information: Genetic history of North Africa and Proto-Berber language
Ancient Libu Libyan. Bronze inlaid with gold and silver, during the reign of Rameses II, Louvre Museum.
The prehistoric populations of North Africa are related to the wider group of Paleo-Mediterranean peoples.
The Afroasiatic family may have originated in the mesolithic period, perhaps in the context of the Capsian culture.[21][22]
DNA analysis has found commonalities between Berber populations and those of the Sami people of Scandinavia showing a link dating from around 9,000 years ago.[23]
By 5000 BC, the populations of North Africa are an amalgamation of Ibero-Maurisian and Capsian stock blended with a more recent intrusion associated with the Neolithic revolution.[24]
Out of these populations, the proto-Berber tribes form during the Late Bronze to Early Iron Age.[25]
Sami people of Scandinavia
http://en.wikipedia.org/wiki/Sami_people
ジブラルタルコースもあり得るようだww
http://en.wikipedia.org/wiki/Berber_people
Origins[edit]
Further information: Genetic history of North Africa and Proto-Berber language
Ancient Libu Libyan. Bronze inlaid with gold and silver, during the reign of Rameses II, Louvre Museum.
The prehistoric populations of North Africa are related to the wider group of Paleo-Mediterranean peoples.
The Afroasiatic family may have originated in the mesolithic period, perhaps in the context of the Capsian culture.[21][22]
DNA analysis has found commonalities between Berber populations and those of the Sami people of Scandinavia showing a link dating from around 9,000 years ago.[23]
By 5000 BC, the populations of North Africa are an amalgamation of Ibero-Maurisian and Capsian stock blended with a more recent intrusion associated with the Neolithic revolution.[24]
Out of these populations, the proto-Berber tribes form during the Late Bronze to Early Iron Age.[25]
Sami people of Scandinavia
http://en.wikipedia.org/wiki/Sami_people
>>704
http://en.wikipedia.org/wiki/Sami_history
Origin[edit]
The Sami are claimed by some to be the aboriginal Northern Europeans, possibly those who first reentered Europe from ice age refugia after the last glacial maximum.
The genetic origin of the Sami is still unknown, though recent genetic research may be providing some clues.
Nevertheless, it appears that the Sami represent an old Asian and European population.
Archeological evidence suggests that people along the southern shores of Lake Onega and around Lake Ladoga reached the River Utsjoki in Northern Finnish Lapland before 8100 BC.[7]
However, it is not likely that Sami languages are so old.
According to the comparative linguist Ante Aikio, the Sami proto-language developed in South Finland or in Karelia around 2000?2500 years ago, spreading then to northern Fennoscandia.[8]
In any case, the Sami are the earliest of the contemporary ethnic groups represented in the Sami area, and are consequently considered the indigenous population of the area.[citation
The genetic lineage of the Sami is unique, and may reflect an early history of geographic isolation, genetic drift, and genetic bottle-necking.
The uniqueness of the Sami gene pool has made it one of the most extensively studied genetic population in the world.
The most frequent Sami MtDNA (female) haplotype is U5b1, with type V also common.[citation needed]
Ice age
Stone Age
も見ておきたいかな
http://en.wikipedia.org/wiki/Sami_history
Origin[edit]
The Sami are claimed by some to be the aboriginal Northern Europeans, possibly those who first reentered Europe from ice age refugia after the last glacial maximum.
The genetic origin of the Sami is still unknown, though recent genetic research may be providing some clues.
Nevertheless, it appears that the Sami represent an old Asian and European population.
Archeological evidence suggests that people along the southern shores of Lake Onega and around Lake Ladoga reached the River Utsjoki in Northern Finnish Lapland before 8100 BC.[7]
However, it is not likely that Sami languages are so old.
According to the comparative linguist Ante Aikio, the Sami proto-language developed in South Finland or in Karelia around 2000?2500 years ago, spreading then to northern Fennoscandia.[8]
In any case, the Sami are the earliest of the contemporary ethnic groups represented in the Sami area, and are consequently considered the indigenous population of the area.[citation
The genetic lineage of the Sami is unique, and may reflect an early history of geographic isolation, genetic drift, and genetic bottle-necking.
The uniqueness of the Sami gene pool has made it one of the most extensively studied genetic population in the world.
The most frequent Sami MtDNA (female) haplotype is U5b1, with type V also common.[citation needed]
Ice age
Stone Age
も見ておきたいかな
ムーア人
http://ja.wikipedia.org/wiki/%E3%83%A0%E3%83%BC%E3%82%A2%E4%BA%BA
ムーア人(英: Moors)は、北西アフリカのイスラム教教徒の呼称。主にベルベル人を指して用いられる。
ムーア人の横顔[編集]はちまきを巻いた『ムーア人の横顔』は、フランスのコルシカ島とイタリアのサルデーニャ島の旗になっている。
コルシカ島の島旗にムーア人の横顔を採用したのは、コルシカの英雄パスカル・パオリである[1]。
http://ja.wikipedia.org/wiki/%E3%83%A0%E3%83%BC%E3%82%A2%E4%BA%BA
ムーア人(英: Moors)は、北西アフリカのイスラム教教徒の呼称。主にベルベル人を指して用いられる。
ムーア人の横顔[編集]はちまきを巻いた『ムーア人の横顔』は、フランスのコルシカ島とイタリアのサルデーニャ島の旗になっている。
コルシカ島の島旗にムーア人の横顔を採用したのは、コルシカの英雄パスカル・パオリである[1]。
ノルマンコンクエストやイスラーム
の影響はあんまり見えてこないんだよな
の影響はあんまり見えてこないんだよな
盛りだくさんでワラタ
サルデーニャ
http://ja.wikipedia.org/wiki/%E3%82%B5%E3%83%AB%E3%83%87%E3%83%BC%E3%83%8B%E3%83%A3
歴史[編集]
新石器時代からローマ帝国の時代にかけて、ヌラーゲ(Nuraghe)人が上陸し、生活しはじめた。
現在もこの民族の子孫が7000人以上いる。この謎に満ちた民族は、紀元前20世紀頃、東地中海からやって来たものと推測されている。
少しわかっていることは、エジプトの碑文に「海の民」という意味の名前で登場する人々を指しているということである。
その碑文の研究によると、彼らは、サルディス(リディア)を出発し、ティレニア海にたどり着いた。そこで、サルデーニャに行く者とエトルリアに行く者に分かれた、ということである。
しかし、サルデーニャ人の起源に関する理論のほとんどは、遺伝学的な研究と民族の移動状況を重要視している。
遺伝学的な研究によると、サルデーニャ人は周辺地域の人々や若い民族とは異なり、前インド=ヨーロッパ人だとしている。
新石器時代以降の遺跡の散らばりぐあい、点在範囲、その大きさを調べれば、島の大体の人口がわかり、また彼らがこの島のどこに上陸し、定着したかがわかる。
以下は英語版の項目en:History of Sardiniaからの翻訳である。
先史時代[編集]1979年、15万年前にさかのぼる人類の痕跡が発見された。
ガッルーラからサルデーニャ北部に居住した最初の人間は、おそらくイタリア半島のトスカーナから渡ってきたとみられている。
島の中央部にはバレアレス海を渡り、イベリア半島から来た人々が居住したとも考えられている。
先史時代の矢じり(約5000年?6000年前)や、現在カリャリの考古学博物館に納められている地中海地方の母神像から、高いレベルで石の彫刻を作る能力を持っていたと推測される。
サルデーニャ
http://ja.wikipedia.org/wiki/%E3%82%B5%E3%83%AB%E3%83%87%E3%83%BC%E3%83%8B%E3%83%A3
歴史[編集]
新石器時代からローマ帝国の時代にかけて、ヌラーゲ(Nuraghe)人が上陸し、生活しはじめた。
現在もこの民族の子孫が7000人以上いる。この謎に満ちた民族は、紀元前20世紀頃、東地中海からやって来たものと推測されている。
少しわかっていることは、エジプトの碑文に「海の民」という意味の名前で登場する人々を指しているということである。
その碑文の研究によると、彼らは、サルディス(リディア)を出発し、ティレニア海にたどり着いた。そこで、サルデーニャに行く者とエトルリアに行く者に分かれた、ということである。
しかし、サルデーニャ人の起源に関する理論のほとんどは、遺伝学的な研究と民族の移動状況を重要視している。
遺伝学的な研究によると、サルデーニャ人は周辺地域の人々や若い民族とは異なり、前インド=ヨーロッパ人だとしている。
新石器時代以降の遺跡の散らばりぐあい、点在範囲、その大きさを調べれば、島の大体の人口がわかり、また彼らがこの島のどこに上陸し、定着したかがわかる。
以下は英語版の項目en:History of Sardiniaからの翻訳である。
先史時代[編集]1979年、15万年前にさかのぼる人類の痕跡が発見された。
ガッルーラからサルデーニャ北部に居住した最初の人間は、おそらくイタリア半島のトスカーナから渡ってきたとみられている。
島の中央部にはバレアレス海を渡り、イベリア半島から来た人々が居住したとも考えられている。
先史時代の矢じり(約5000年?6000年前)や、現在カリャリの考古学博物館に納められている地中海地方の母神像から、高いレベルで石の彫刻を作る能力を持っていたと推測される。
>>709
ヌラーゲ文化の時代
先史時代のサルデーニャは、ヌラーゲと呼ばれる独特の石造りの構造物に特徴づけられている。
サルデーニャには複雑な構造のものから単純なものまで、大小7000のヌラーゲが現存している。
最も有名なのはカリャリ県バルーミニのヌラーゲ遺跡、スー・ヌラージ・ディ・バルーミニである。
このヌラーゲは紀元前1800年から250年頃にわたって造られ、紀元前1200年から900年頃に全盛期を迎えた。
聖なる水場の隣に建てられ(例:Santa Cristina, Sardara)、墓の構造はドルメンと呼ばれる。
この時代サルデーニャ人は既に、西地中海で交易を行っていたミケーネ人と接触していたことがわかっている。
エジプトを侵略した海洋民族シャルダナ (Shardana) とサルデーニャとのつながりは真偽が疑わしく、立証されていない。
墓場 (Tombe dei giganti) には沈みかけの船をかたどった墓石があり、長い航海中に惨事があったことを示している。
古代ギリシャで初めて地中海を西に航海したエウボイア人は、サルデーニャをHyknousaと呼んだ。
のちにラテン化しIchnus(s)a(イクヌーザ)となった。
ノーラ遺跡の石碑は、フェニキア人がこの島をShardenと呼んだ証拠となっており、これがSardiniaという名前の由来となっている。
サルデーニャにおけるフェニキア人、カルタゴ人、そしてローマ人
紀元前8世紀から、Tharros(ターロス)、Bithia(ビティア)、Sulcis(スルシス)、Nora(ノーラ)、Karalis(カラリス、現在のカリャリ)と、フェニキア人が都市や砦をいくつもサルデーニャに築いた。
フェニキア人はレバノンの出身で、地中海で交易を行っていた。彼らは島のあらゆるエリアに定住した。
サルデーニャはカルタゴ(現在のチュニジア)、スペイン、ローヌ川(フランス)、エトルリア(イタリア半島)の間にあったため、西地中海の中心として特別な地位を獲得していた。
イグレージアス周辺の鉱物地帯は、鉛や亜鉛の産地として重要であった。
都市は防御しやすく天然の港になる、多くは河口に近い半島部や島のような、戦略上の重要な地点に造られた。
フェニキア人ののちに、紀元前500年ごろカルタゴ人(Punic、ポエニ)がサルデーニャ周辺の地中海の覇権を確立した。
カルタゴの影響はサルデーニャのほぼ全域に及んでいる。
ヌラーゲ文化の時代
先史時代のサルデーニャは、ヌラーゲと呼ばれる独特の石造りの構造物に特徴づけられている。
サルデーニャには複雑な構造のものから単純なものまで、大小7000のヌラーゲが現存している。
最も有名なのはカリャリ県バルーミニのヌラーゲ遺跡、スー・ヌラージ・ディ・バルーミニである。
このヌラーゲは紀元前1800年から250年頃にわたって造られ、紀元前1200年から900年頃に全盛期を迎えた。
聖なる水場の隣に建てられ(例:Santa Cristina, Sardara)、墓の構造はドルメンと呼ばれる。
この時代サルデーニャ人は既に、西地中海で交易を行っていたミケーネ人と接触していたことがわかっている。
エジプトを侵略した海洋民族シャルダナ (Shardana) とサルデーニャとのつながりは真偽が疑わしく、立証されていない。
墓場 (Tombe dei giganti) には沈みかけの船をかたどった墓石があり、長い航海中に惨事があったことを示している。
古代ギリシャで初めて地中海を西に航海したエウボイア人は、サルデーニャをHyknousaと呼んだ。
のちにラテン化しIchnus(s)a(イクヌーザ)となった。
ノーラ遺跡の石碑は、フェニキア人がこの島をShardenと呼んだ証拠となっており、これがSardiniaという名前の由来となっている。
サルデーニャにおけるフェニキア人、カルタゴ人、そしてローマ人
紀元前8世紀から、Tharros(ターロス)、Bithia(ビティア)、Sulcis(スルシス)、Nora(ノーラ)、Karalis(カラリス、現在のカリャリ)と、フェニキア人が都市や砦をいくつもサルデーニャに築いた。
フェニキア人はレバノンの出身で、地中海で交易を行っていた。彼らは島のあらゆるエリアに定住した。
サルデーニャはカルタゴ(現在のチュニジア)、スペイン、ローヌ川(フランス)、エトルリア(イタリア半島)の間にあったため、西地中海の中心として特別な地位を獲得していた。
イグレージアス周辺の鉱物地帯は、鉛や亜鉛の産地として重要であった。
都市は防御しやすく天然の港になる、多くは河口に近い半島部や島のような、戦略上の重要な地点に造られた。
フェニキア人ののちに、紀元前500年ごろカルタゴ人(Punic、ポエニ)がサルデーニャ周辺の地中海の覇権を確立した。
カルタゴの影響はサルデーニャのほぼ全域に及んでいる。
>>704>>706
Saami and Berbers?An Unexpected Mitochondrial DNA Link
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1199377/
なんでmtDNAだけだね
Seven of the new sequences (one Berber from Algeria, two Italian, one Spanish, and three Saami) clustered into U5b1b,
the subclade encompassing the Yakut and Fulbe mtDNAs.
The Saami and the Yakut mtDNAs formed a minor branch distinguished only by the transition at nt 16144,
the Berber and the Fulbe mtDNAs clustered in a second minor branch also characterized only by control-region mutations,
and the Italian and Spanish mtDNAs formed other minor branches.
ちなみにU5b1bは>>692に近縁者があまり居ないね
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1199377/figure/FG1/
にはUの系統が詳しく載ってるからなにかの参考に
Saami and Berbers?An Unexpected Mitochondrial DNA Link
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1199377/
なんでmtDNAだけだね
Seven of the new sequences (one Berber from Algeria, two Italian, one Spanish, and three Saami) clustered into U5b1b,
the subclade encompassing the Yakut and Fulbe mtDNAs.
The Saami and the Yakut mtDNAs formed a minor branch distinguished only by the transition at nt 16144,
the Berber and the Fulbe mtDNAs clustered in a second minor branch also characterized only by control-region mutations,
and the Italian and Spanish mtDNAs formed other minor branches.
ちなみにU5b1bは>>692に近縁者があまり居ないね
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1199377/figure/FG1/
にはUの系統が詳しく載ってるからなにかの参考に
713 :世界@名無史さん:2014/01/14(火) 01:20:49.76 0
>>706 さらにはサーミは広顔短頭で、少なくとも後者に関しては現生人類の祖先的形質とは言い難いしな
まあY-DNAでは北アジア系のNが多いようなので、北欧先住民と北方モンゴロイドの融合した子孫なんだろうけど
>>709,710 「ヌラーゲ人の子孫が今でも7000人以上」というのはおそらく「ヌラーゲと言う構造物が7000以上」の間違いだろうな
そもそも文化が絶えて数千年後に、どうやってヌラーゲ人と他のサルデーニャ人を区別するんだw
まあY-DNAでは北アジア系のNが多いようなので、北欧先住民と北方モンゴロイドの融合した子孫なんだろうけど
>>709,710 「ヌラーゲ人の子孫が今でも7000人以上」というのはおそらく「ヌラーゲと言う構造物が7000以上」の間違いだろうな
そもそも文化が絶えて数千年後に、どうやってヌラーゲ人と他のサルデーニャ人を区別するんだw
>>713
うーん、解釈が間違ってなければ
ここ15,000年程度の間にイベリアの北端を挟んで
遺伝的交流(女性が移動して入植)があって
それもおそらくは7600年より
最近であるという話なんじゃないかと思うんだが。
>>693>>695
そんでその頃にイベリア北端にいた>>695>>700の
(WHG) West European Hunter-Gatherers は
青い目をしていたという話
まぁ、俺は英語読めないからホントのとこ、
どうなのかってのがあるんだけどねww
うーん、解釈が間違ってなければ
ここ15,000年程度の間にイベリアの北端を挟んで
遺伝的交流(女性が移動して入植)があって
それもおそらくは7600年より
最近であるという話なんじゃないかと思うんだが。
>>693>>695
そんでその頃にイベリア北端にいた>>695>>700の
(WHG) West European Hunter-Gatherers は
青い目をしていたという話
まぁ、俺は英語読めないからホントのとこ、
どうなのかってのがあるんだけどねww
715 :世界@名無史さん:2014/01/15(水) 17:03:27.34 0
>>701
北欧も主なY遺伝子ハプロタイプだけでも、I1、R1a、R1b、N1C1とあるからね。
さらに紀元前には中近東系の遺伝子(J2、E?)も少なからず存在したようだから。
簡単には言いきれないだろうね。
北欧も主なY遺伝子ハプロタイプだけでも、I1、R1a、R1b、N1C1とあるからね。
さらに紀元前には中近東系の遺伝子(J2、E?)も少なからず存在したようだから。
簡単には言いきれないだろうね。
>>711
俺の適当な記憶によると
サルディーニャはフェニキアの遠方航海の前線基地だったような気がして、
遠航海するときは各地に殖民農地を築いて補給をしたような気がして
北はアイルランドで南は紅海まで船を漕いだ気がしている
俺の適当な記憶によると
サルディーニャはフェニキアの遠方航海の前線基地だったような気がして、
遠航海するときは各地に殖民農地を築いて補給をしたような気がして
北はアイルランドで南は紅海まで船を漕いだ気がしている
>>710
Nora, Italy
http://en.wikipedia.org/wiki/Nora,_Italy
According to legend, Nora was founded by a group of Iberians from Tartessus led by Norax, a mythological hero son of Eriteide and the god Hermes.
It is believed to be the first town founded in Sardinia and to have been settled by the ancient Sherden or the Nuraghi people, and later colonized by Phoenicians.
Ancient sources[edit]
According with Pausanias, Norace came in Sardinia at the helm of the Iberians who later founded the city of Nora.[1]
Solinus specifically stated that Norace arrived in Sardinia from the mythical city of Tartessos located in southern Iberia.[2]
Tharros
http://en.wikipedia.org/wiki/Tharros
Tharros (also spelled Tharras, Greek: Θ?ρρα?, Ptol., Tarrae or Tarras) was an ancient city on the west coast of Sardinia, Italy
Archaeological research done in the area of Tharros has established that in the eighth century BC the town was founded by Phoenicians.
On the remains of a former village built by the nuragic peoples (1900-730 BC[1]) on top of the hill called Su Muru Mannu they founded a tophet,
an open air sacred place common for several installations of Phoenicians in the western Mediterranean, and seen as a first sign of colonization and urbanization
Nora, Italy
http://en.wikipedia.org/wiki/Nora,_Italy
According to legend, Nora was founded by a group of Iberians from Tartessus led by Norax, a mythological hero son of Eriteide and the god Hermes.
It is believed to be the first town founded in Sardinia and to have been settled by the ancient Sherden or the Nuraghi people, and later colonized by Phoenicians.
Ancient sources[edit]
According with Pausanias, Norace came in Sardinia at the helm of the Iberians who later founded the city of Nora.[1]
Solinus specifically stated that Norace arrived in Sardinia from the mythical city of Tartessos located in southern Iberia.[2]
Tharros
http://en.wikipedia.org/wiki/Tharros
Tharros (also spelled Tharras, Greek: Θ?ρρα?, Ptol., Tarrae or Tarras) was an ancient city on the west coast of Sardinia, Italy
Archaeological research done in the area of Tharros has established that in the eighth century BC the town was founded by Phoenicians.
On the remains of a former village built by the nuragic peoples (1900-730 BC[1]) on top of the hill called Su Muru Mannu they founded a tophet,
an open air sacred place common for several installations of Phoenicians in the western Mediterranean, and seen as a first sign of colonization and urbanization
>>710
Cagliari
http://en.wikipedia.org/wiki/Cagliari
Early history[edit]
Cagliari has been inhabited since ancient times.
It occupies a favourable position between the sea and a fertile plain, and is surrounded by two swamps (which increased the defense from inner lands)
and is close to high mountains towards which people could evacuate if everything else was lost.
Some findings of prehistoric inhabitants were found in Monte Claro (the eponymous culture, from the Copper Age) and in Cape Sant'Elia (several domus de janas).
Sulci
http://en.wikipedia.org/wiki/Sulci
The foundation of Sulci is expressly attributed to the Carthaginians (Paus. x. 17. § 9; Claudian, B. Gild. 518),
and it seems to have become under that people one of the most considerable cities of Sardinia, and one of the chief seats of their power in the island.
ちなみに俺のサルディーニャのイメージは
羊飼いが人攫ってる国
Cagliari
http://en.wikipedia.org/wiki/Cagliari
Early history[edit]
Cagliari has been inhabited since ancient times.
It occupies a favourable position between the sea and a fertile plain, and is surrounded by two swamps (which increased the defense from inner lands)
and is close to high mountains towards which people could evacuate if everything else was lost.
Some findings of prehistoric inhabitants were found in Monte Claro (the eponymous culture, from the Copper Age) and in Cape Sant'Elia (several domus de janas).
Sulci
http://en.wikipedia.org/wiki/Sulci
The foundation of Sulci is expressly attributed to the Carthaginians (Paus. x. 17. § 9; Claudian, B. Gild. 518),
and it seems to have become under that people one of the most considerable cities of Sardinia, and one of the chief seats of their power in the island.
ちなみに俺のサルディーニャのイメージは
羊飼いが人攫ってる国
>>690
アゾレス諸島と勘違いしたかの、、、
大西洋のアゾレス諸島でカルタゴ人の遺跡を発見
http://www.archeonews.net/2011/07/blog-post_13.html
ポルトガル考古学調査協会(Portuguese Association of Archeological Research:略称APIA)は、
ポルトガル領のアゾレス諸島において、紀元前4世紀以後のカルタゴ人による女神タニトの神殿を発見した。
アゾレス諸島はポルトガル沖約1000kmの大西洋上にある。
1427年にポルトガル人によって発見される前のアゾレス諸島における人類の足跡は、これまであまり知られていなかった。
遺跡はテルセイラ島のモンテ・ブラジル、アングラ・ド・エロイズモで発見された。
APIAの考古学者Nuno Ribeiro氏とAnabela Joaquinito氏によれば、5つ以上の地下神殿と、少なうとも3つの岩を掘ってつくられた聖域が発見されている。
発見された遺構には岩に掘り込まれた水槽があり、水をささげるための水路につながっていた。
岩に掘り込まれた椅子や、植物に覆われた儀式的な槽、多くの柱穴が残っていた。
柱穴はこの場所に覆い屋があったことを示している。
APIAの調査隊によれば、1つ目の神殿には4つの水槽があって、儀式での献水に使用する新鮮な水を集めるための水路へとつながっているという。
献水は犠牲の儀式に使用された可能性が高いと考えられている。
2つ目と3つ目の神殿はフォルテ・デ・サン=ディオゴという地域に位置しており、先月6月に発見されていた。
最近では、イスラム教スンニ派によって書かれた文字がサン・ミゲル島とテルセイラ島で発見された。
その前にはカルタゴとギリシアの多くの硬貨がコルヴォ島から発見されており、また多くの先キリスト教時代の地下構造物がテルセイラ島、コルヴォ島から見つかっていた。
同様の発見はサンタ・マリア島やフローレス島でも報告されている。
これらの発見から、15世紀のポルトガルによる占領以前に、人類がアゾレス諸島で活動していたということが、はっきりと言えるようになってきている。
アゾレス諸島と勘違いしたかの、、、
大西洋のアゾレス諸島でカルタゴ人の遺跡を発見
http://www.archeonews.net/2011/07/blog-post_13.html
ポルトガル考古学調査協会(Portuguese Association of Archeological Research:略称APIA)は、
ポルトガル領のアゾレス諸島において、紀元前4世紀以後のカルタゴ人による女神タニトの神殿を発見した。
アゾレス諸島はポルトガル沖約1000kmの大西洋上にある。
1427年にポルトガル人によって発見される前のアゾレス諸島における人類の足跡は、これまであまり知られていなかった。
遺跡はテルセイラ島のモンテ・ブラジル、アングラ・ド・エロイズモで発見された。
APIAの考古学者Nuno Ribeiro氏とAnabela Joaquinito氏によれば、5つ以上の地下神殿と、少なうとも3つの岩を掘ってつくられた聖域が発見されている。
発見された遺構には岩に掘り込まれた水槽があり、水をささげるための水路につながっていた。
岩に掘り込まれた椅子や、植物に覆われた儀式的な槽、多くの柱穴が残っていた。
柱穴はこの場所に覆い屋があったことを示している。
APIAの調査隊によれば、1つ目の神殿には4つの水槽があって、儀式での献水に使用する新鮮な水を集めるための水路へとつながっているという。
献水は犠牲の儀式に使用された可能性が高いと考えられている。
2つ目と3つ目の神殿はフォルテ・デ・サン=ディオゴという地域に位置しており、先月6月に発見されていた。
最近では、イスラム教スンニ派によって書かれた文字がサン・ミゲル島とテルセイラ島で発見された。
その前にはカルタゴとギリシアの多くの硬貨がコルヴォ島から発見されており、また多くの先キリスト教時代の地下構造物がテルセイラ島、コルヴォ島から見つかっていた。
同様の発見はサンタ・マリア島やフローレス島でも報告されている。
これらの発見から、15世紀のポルトガルによる占領以前に、人類がアゾレス諸島で活動していたということが、はっきりと言えるようになってきている。
2011/06/08
ヨーロッパ最古? 7000年前の都市がブルガリアで発見される
http://www.archeonews.net/2011/06/7000.html
ブルガリアで、ヨーロッパ最古と考えられる町が発見された、と地元メディアが報じている。
遺跡はブルガリア南西部にあるパザルジク市の近くにある。
2008年には考古学者のYasen Boyadzhiev氏が率いるチームが、この地域で広大な墓地を発見していた。
墓地はこの地域の名前であるYunatsiteとして知られるようになった。
その後調査区を拡大した結果、面積10万平方メートルにおよぶ紀元前4700年〜4600年頃の巨大な都市が先日発見された。
Boyadzhiev氏によると、発見された遺跡は都市センターとしての全ての特徴を備えているという。
また、一部では幅5m、高さ5mもある城壁、溝、その他の防御のための壁が発見されている。
城塞は町の中でも高所だけを覆っており、壁の外にも建築物は続いている。
壁の内側には住居だけでなく、明らかに工房街と思われるものも発見された。
発掘された遺物のいくつかは、極めて高い技術で作られていたという。
これまでのところ、これほど大規模で計画的な町の建設は、ずっと後の時代でしか知られていなかった。
「中世のヨーロッパに見られるような都市計画が、中世よりも数千年古い時代にあった」とBoyadzhiev氏は述べている。
ヨーロッパ最古? 7000年前の都市がブルガリアで発見される
http://www.archeonews.net/2011/06/7000.html
ブルガリアで、ヨーロッパ最古と考えられる町が発見された、と地元メディアが報じている。
遺跡はブルガリア南西部にあるパザルジク市の近くにある。
2008年には考古学者のYasen Boyadzhiev氏が率いるチームが、この地域で広大な墓地を発見していた。
墓地はこの地域の名前であるYunatsiteとして知られるようになった。
その後調査区を拡大した結果、面積10万平方メートルにおよぶ紀元前4700年〜4600年頃の巨大な都市が先日発見された。
Boyadzhiev氏によると、発見された遺跡は都市センターとしての全ての特徴を備えているという。
また、一部では幅5m、高さ5mもある城壁、溝、その他の防御のための壁が発見されている。
城塞は町の中でも高所だけを覆っており、壁の外にも建築物は続いている。
壁の内側には住居だけでなく、明らかに工房街と思われるものも発見された。
発掘された遺物のいくつかは、極めて高い技術で作られていたという。
これまでのところ、これほど大規模で計画的な町の建設は、ずっと後の時代でしか知られていなかった。
「中世のヨーロッパに見られるような都市計画が、中世よりも数千年古い時代にあった」とBoyadzhiev氏は述べている。
>>708
大事なのを忘れていた
ヴァンダル族
http://ja.wikipedia.org/wiki/%E3%83%B4%E3%82%A1%E3%83%B3%E3%83%80%E3%83%AB%E6%97%8F
ヴァンダル族(Vandal)は、ローマ帝国末期にヨーロッパ中央部に侵入してきた系統不明の民族で、北アフリカのカルタゴを中心にヴァンダル王国(フランス語版)を建国した。
彼らが北アフリカに進出する前に一時的に定着したスペインのアンダルシア(もともとはVandalusiaと綴った)は、おそらくヴァンダル族に由来する地名である。
また彼らの名前は、ヴァンダリズムの語源ともなっている。
ヴァンダル族は、シリンジイ(Silingii)とハスディンジイ(Hasdingii)の2つの系統に分けられる。
シリンジイは、ゲルマニア・マグナ(いわゆるゲルマニア)のうち、後にシレジアと呼ばれるようになった地域とその周辺に何世紀にも渡って住んでいた。
2世紀に、ハスディンジイは、王のラウスとラプトに率いられて南に移動し、ドナウ川下流でローマ帝国を攻撃しはじめた。
その後、ローマ帝国と和睦しルーマニアの西ダキアとハンガリーに定着した。
406年12月31日、ヴァンダル族は凍結したライン川を渡り、ガリアに侵入した。
ゴディギゼルの息子グンデリク(イタリア語版)王率いるヴァンダル族は、ガリアの西や南へ略奪して回った。
409年10月、ピレネー山脈を越えてスペインに入った。そこは、ローマ帝国から建国を許された土地であった。
アラン族がポルトガルとカルタヘナ一帯を領有し、ヴァンダル族はガリシアとアンダルシアを得た。
ヴァンダル王国(470年)グンデリクの兄弟ゲイセリック(ガイセリック)は、艦隊の建造を始めた。
38歳のゲイセリックが王になった後の429年、ジブラルタル海峡を渡り、アフリカ沿岸をカルタゴに向かって東方に移動しはじめた。
435年に、ローマ帝国は北アフリカのいくつかの領土を彼らに与えたが、439年、ヴァンダル族は自らカルタゴを占領した。
ゲイセリックはここにヴァンダル族とアラン族からなるヴァンダル王国(フランス語版)を建国した。
この王国は地中海における一大勢力となり、シチリア島、サルデニア島、コルシカ島、バレアレス諸島を征服している。
大事なのを忘れていた
ヴァンダル族
http://ja.wikipedia.org/wiki/%E3%83%B4%E3%82%A1%E3%83%B3%E3%83%80%E3%83%AB%E6%97%8F
ヴァンダル族(Vandal)は、ローマ帝国末期にヨーロッパ中央部に侵入してきた系統不明の民族で、北アフリカのカルタゴを中心にヴァンダル王国(フランス語版)を建国した。
彼らが北アフリカに進出する前に一時的に定着したスペインのアンダルシア(もともとはVandalusiaと綴った)は、おそらくヴァンダル族に由来する地名である。
また彼らの名前は、ヴァンダリズムの語源ともなっている。
ヴァンダル族は、シリンジイ(Silingii)とハスディンジイ(Hasdingii)の2つの系統に分けられる。
シリンジイは、ゲルマニア・マグナ(いわゆるゲルマニア)のうち、後にシレジアと呼ばれるようになった地域とその周辺に何世紀にも渡って住んでいた。
2世紀に、ハスディンジイは、王のラウスとラプトに率いられて南に移動し、ドナウ川下流でローマ帝国を攻撃しはじめた。
その後、ローマ帝国と和睦しルーマニアの西ダキアとハンガリーに定着した。
406年12月31日、ヴァンダル族は凍結したライン川を渡り、ガリアに侵入した。
ゴディギゼルの息子グンデリク(イタリア語版)王率いるヴァンダル族は、ガリアの西や南へ略奪して回った。
409年10月、ピレネー山脈を越えてスペインに入った。そこは、ローマ帝国から建国を許された土地であった。
アラン族がポルトガルとカルタヘナ一帯を領有し、ヴァンダル族はガリシアとアンダルシアを得た。
ヴァンダル王国(470年)グンデリクの兄弟ゲイセリック(ガイセリック)は、艦隊の建造を始めた。
38歳のゲイセリックが王になった後の429年、ジブラルタル海峡を渡り、アフリカ沿岸をカルタゴに向かって東方に移動しはじめた。
435年に、ローマ帝国は北アフリカのいくつかの領土を彼らに与えたが、439年、ヴァンダル族は自らカルタゴを占領した。
ゲイセリックはここにヴァンダル族とアラン族からなるヴァンダル王国(フランス語版)を建国した。
この王国は地中海における一大勢力となり、シチリア島、サルデニア島、コルシカ島、バレアレス諸島を征服している。
722 :世界@名無史さん:2014/01/16(木) 17:36:21.66 0
>>718 英語で「冷笑的な」を意味するsardonicは、「サルデーニャの」と言うのが原義だそうだな
隔絶していたようでいていろんな騒乱に巻き込まれて、物の見方が斜に構えたようになったか
隔絶していたようでいていろんな騒乱に巻き込まれて、物の見方が斜に構えたようになったか
723 :世界@名無史さん:2014/01/16(木) 19:06:15.88 0
2030年までにイスラム国家になるベルギー
http://www.youtube.com/watch?v=VdHEAG8DEP4・・・動画/5分40秒/CBN NEWS
パリに出現した移民自治区
http://www.youtube.com/watch?v=w9OyDm_t0yI・・・動画/4分59秒/CBN NEWS
移民問題 ロンドン
http://www.youtube.com/watch?v=0fjztxZ2MZw・・・動画/3分46秒
移民亡国ドイツ
http://www.youtube.com/watch?v=Rvb-7KfKPwA・・・動画/12分56秒
ウェ−デンは2049年までにイスラム国家
http://www.youtube.com/watch?v=UocmQZa8tqo・・・動画/6分
http://www.youtube.com/watch?v=VdHEAG8DEP4・・・動画/5分40秒/CBN NEWS
パリに出現した移民自治区
http://www.youtube.com/watch?v=w9OyDm_t0yI・・・動画/4分59秒/CBN NEWS
移民問題 ロンドン
http://www.youtube.com/watch?v=0fjztxZ2MZw・・・動画/3分46秒
移民亡国ドイツ
http://www.youtube.com/watch?v=Rvb-7KfKPwA・・・動画/12分56秒
ウェ−デンは2049年までにイスラム国家
http://www.youtube.com/watch?v=UocmQZa8tqo・・・動画/6分
>>720
http://www.afpbb.com/articles/-/2910148?pid=9750304
【11月1日 AFP】ブルガリア東部・黒海沿岸の都市バルナ(Varna)近郊で、欧州最古とみられる先史時代の「町」の跡と、塩の生産設備が発掘された。
当時、塩は希少品だったことから、考古学者らはバルナにある古代墓地で大量の金の装飾品が見つかる理由がこれで解明できると期待している。
「町」が発掘されたのは、バルナ郊外の町プロバディヤ(Provadia)近郊にあるプロバディヤ・サルニツァタ(Solnitsata)遺跡。
2005年から続けられてきた発掘調査で、2階建て住宅や墓地、儀式に使われたとみられる多数の穴、門の一部、要塞らしき建造物や防壁が発見された。
炭素年代測定で、全て紀元前4700〜4200年の「金石併用時代」中期〜後期のものであることが分かったという。
ブルガリア国立考古学研究所(National Institute of Archeology)の考古学チームは、墓地に埋葬されていた人骨の姿勢や出土した埋葬品がいずれも、
ブルガリアで過去に見つかった新石器時代のものとは大きく異なっている点を指摘。
また、「町」を取り囲む石造りの巨大な防壁も、これまで南東欧で見つかった先史時代の遺跡には見られなかったものだとして、「大変興味深い発見だ」と説明している。
人口350人ほどだったとみられるこの集落は防壁でしっかりと守られ、宗教の中心地だったと考えられる。
さらに最も重要な点として、広く交易に使われていた塩を特産品としていた。以上から研究チームは、
欧州で知られている中で最古の「先史時代の町」と呼ぶのにふさわしい全ての条件を備えていると結論付けた。
バルナには紀元前4300年ごろにさかのぼるネクロポリス(古代文明の集団墓地)があり、世界最古とされる黄金の装飾品が多数発掘されている。
しかし、農業や牧畜が中心で資源も少ないこの地域に、なぜこれほどの富が集中したのかは謎だった。
研究チームでは、塩の交易こそが富をもたらしたのではないかと考えている。
(c)AFP/Diana SIMEONOVA
http://www.afpbb.com/articles/-/2910148?pid=9750304
【11月1日 AFP】ブルガリア東部・黒海沿岸の都市バルナ(Varna)近郊で、欧州最古とみられる先史時代の「町」の跡と、塩の生産設備が発掘された。
当時、塩は希少品だったことから、考古学者らはバルナにある古代墓地で大量の金の装飾品が見つかる理由がこれで解明できると期待している。
「町」が発掘されたのは、バルナ郊外の町プロバディヤ(Provadia)近郊にあるプロバディヤ・サルニツァタ(Solnitsata)遺跡。
2005年から続けられてきた発掘調査で、2階建て住宅や墓地、儀式に使われたとみられる多数の穴、門の一部、要塞らしき建造物や防壁が発見された。
炭素年代測定で、全て紀元前4700〜4200年の「金石併用時代」中期〜後期のものであることが分かったという。
ブルガリア国立考古学研究所(National Institute of Archeology)の考古学チームは、墓地に埋葬されていた人骨の姿勢や出土した埋葬品がいずれも、
ブルガリアで過去に見つかった新石器時代のものとは大きく異なっている点を指摘。
また、「町」を取り囲む石造りの巨大な防壁も、これまで南東欧で見つかった先史時代の遺跡には見られなかったものだとして、「大変興味深い発見だ」と説明している。
人口350人ほどだったとみられるこの集落は防壁でしっかりと守られ、宗教の中心地だったと考えられる。
さらに最も重要な点として、広く交易に使われていた塩を特産品としていた。以上から研究チームは、
欧州で知られている中で最古の「先史時代の町」と呼ぶのにふさわしい全ての条件を備えていると結論付けた。
バルナには紀元前4300年ごろにさかのぼるネクロポリス(古代文明の集団墓地)があり、世界最古とされる黄金の装飾品が多数発掘されている。
しかし、農業や牧畜が中心で資源も少ないこの地域に、なぜこれほどの富が集中したのかは謎だった。
研究チームでは、塩の交易こそが富をもたらしたのではないかと考えている。
(c)AFP/Diana SIMEONOVA
>>720
http://en.wikipedia.org/wiki/Varna_Necropolis
The Varna Necropolis (Bulgarian: Варненски некропол) (also Varna Cemetery) is a burial site in the western industrial zone
of Varna (approximately half a kilometre from Lake Varna and 4 km from the city centre), Bulgaria,
internationally considered one of the key archaeological sites in world prehistory.
The oldest golden treasure in the world, dating from 4,600 BC to 4,200 BC, was discovered at the site.[1]
http://en.wikipedia.org/wiki/Lake_Varna
http://en.wikipedia.org/wiki/File:Lake-Varna-map-EN.gif
http://en.wikipedia.org/wiki/Varna_Necropolis
The Varna Necropolis (Bulgarian: Варненски некропол) (also Varna Cemetery) is a burial site in the western industrial zone
of Varna (approximately half a kilometre from Lake Varna and 4 km from the city centre), Bulgaria,
internationally considered one of the key archaeological sites in world prehistory.
The oldest golden treasure in the world, dating from 4,600 BC to 4,200 BC, was discovered at the site.[1]
http://en.wikipedia.org/wiki/Lake_Varna
http://en.wikipedia.org/wiki/File:Lake-Varna-map-EN.gif
{kind=link}
>>724
http://en.wikipedia.org/wiki/Solnitsata
Salt production drove Solnitsata's economy, and the town is believed to have supplied salt throughout the Balkans.
A large collection of gold objects nearby has led archaeologists to speculate that this trade resulted in considerable wealth for the town's residents.[1]
The town is believed to have been destroyed by an earthquake.[4]
まぁ、この頃には結構な交易網があったってこったね。
時代的には銅の時代だけれども
http://en.wikipedia.org/wiki/Solnitsata
Salt production drove Solnitsata's economy, and the town is believed to have supplied salt throughout the Balkans.
A large collection of gold objects nearby has led archaeologists to speculate that this trade resulted in considerable wealth for the town's residents.[1]
The town is believed to have been destroyed by an earthquake.[4]
まぁ、この頃には結構な交易網があったってこったね。
時代的には銅の時代だけれども
http://www.geocities.jp/e_kamasai/byroad/page12.html
冶金の欧州独自起源説 2012/10/23
この論文(要約)からすると、西アジアよりヨーロッパの方が冶金の始まりは早かったということになりそうだ。
「On the origins of extractive metallurgy: new evidence from Europe」
Abstract
The beginnings of extractive metallurgy in Eurasia are contentious.
The first cast copper objects in this region emerge c. 7000 years ago, and their production has been tentatively linked to centres in the Near East. This assumption,
however, is not substantiated by evidence for copper smelting in those centres.
Here, we present results from recent excavations from Belovode, a Vin?a culture site in Eastern Serbia,
which has provided the earliest direct evidence for copper smelting to date.
The earliest copper smelting activities there took place c. 7000 years ago, contemporary with the emergence of the first cast copper objects.
Through optical, chemical and provenance analyses of copper slag, minerals, ores and artefacts,
we demonstrate the presence of an established metallurgical technology during this period, exploiting multiple sources for raw materials.
These results extend the known record of copper smelting by more than half a millennium, with substantial implications.
Extractive metallurgy occurs at a location far away from the Near East,
challenging the traditional model of a single origin of metallurgy and reviving the possibility of multiple, independent inventions.
(Journal of Archaeological Science Vol.37 (2010) p.2775-2787)
冶金の欧州独自起源説 2012/10/23
この論文(要約)からすると、西アジアよりヨーロッパの方が冶金の始まりは早かったということになりそうだ。
「On the origins of extractive metallurgy: new evidence from Europe」
Abstract
The beginnings of extractive metallurgy in Eurasia are contentious.
The first cast copper objects in this region emerge c. 7000 years ago, and their production has been tentatively linked to centres in the Near East. This assumption,
however, is not substantiated by evidence for copper smelting in those centres.
Here, we present results from recent excavations from Belovode, a Vin?a culture site in Eastern Serbia,
which has provided the earliest direct evidence for copper smelting to date.
The earliest copper smelting activities there took place c. 7000 years ago, contemporary with the emergence of the first cast copper objects.
Through optical, chemical and provenance analyses of copper slag, minerals, ores and artefacts,
we demonstrate the presence of an established metallurgical technology during this period, exploiting multiple sources for raw materials.
These results extend the known record of copper smelting by more than half a millennium, with substantial implications.
Extractive metallurgy occurs at a location far away from the Near East,
challenging the traditional model of a single origin of metallurgy and reviving the possibility of multiple, independent inventions.
(Journal of Archaeological Science Vol.37 (2010) p.2775-2787)
>>726
http://adsabs.harvard.edu/abs/2010EGUGA..12.6393R
Abstract
Recent archeological excavations and findings helped a lot to reconstruct data and information about ancient earthquakes and their effects to the historical societies.
Two cases are under investigations: 4550 years BC Solnitsata-Provadia archaeological site and Cybele temple (6th century BC) site.
The first case is related to the destruction of the defensive bastions of the ancient society of the salt producing ancient civilization.
After field observations and modeling the source of the seismic force is located and the power of the ancient earthquake reconstructed.
The second case is much more complicated but as well as much more informative about the complex disastrous event affected the 8 centuries active temple of the goddess Cybele.
Effects of earthquake, tsunami and landslide activity are well preserved and documented in the soil layers covered the ancient ruins.
Almost all parameters of these hazards are reconstructed by a logic tree, field investigation and documentations and time dependent scenario.
Such investigations are able to help a lot the time frequency of such hazards, but need active participation of the different kinds of experts during the archaeological excavations.
でました Cybele temple(6th century BC)
4550 years BC Solnitsata-Provadia archaeological site
地震、津波、に地すべり
http://adsabs.harvard.edu/abs/2010EGUGA..12.6393R
Abstract
Recent archeological excavations and findings helped a lot to reconstruct data and information about ancient earthquakes and their effects to the historical societies.
Two cases are under investigations: 4550 years BC Solnitsata-Provadia archaeological site and Cybele temple (6th century BC) site.
The first case is related to the destruction of the defensive bastions of the ancient society of the salt producing ancient civilization.
After field observations and modeling the source of the seismic force is located and the power of the ancient earthquake reconstructed.
The second case is much more complicated but as well as much more informative about the complex disastrous event affected the 8 centuries active temple of the goddess Cybele.
Effects of earthquake, tsunami and landslide activity are well preserved and documented in the soil layers covered the ancient ruins.
Almost all parameters of these hazards are reconstructed by a logic tree, field investigation and documentations and time dependent scenario.
Such investigations are able to help a lot the time frequency of such hazards, but need active participation of the different kinds of experts during the archaeological excavations.
でました Cybele temple(6th century BC)
4550 years BC Solnitsata-Provadia archaeological site
地震、津波、に地すべり
>>725
http://en.wikipedia.org/wiki/Durankulak
History[edit]
The Durankulak settlement commenced on a small island, approximately 5200 BC and lasted for thousand years.
The first inhabitants were the Hamangia culture, dated from the middle of the 6th millennium to the middle of 5th millennium BC,
and were the first manifestation of the Neolithic life in Dobruzha.
Hamangia people were small-scale cultivators and plant collectors who built houses, made pottery, herded and hunted animals
Durankulak is the north-easternmost inhabited place in Bulgaria and the northernmost village of the Bulgarian Black Sea Coast,
although the village itself is slightly inland. Durankulak lies north of the town of Shabla,
with the only places to the north along the coast being the formerly exclusively Czechoslovak camping site Kosmos and the Kartalburun and Sivriburun headlands.
Durankulak is also the name of the nearby border checkpoint on the Bulgarian-Romanian border; just north of the border is the Romanian seaside resort Vama Veche.
http://en.wikipedia.org/wiki/Hamangia_culture
The Thinker of Hamangia, Neolithic Hamangia culture (c. 5250-4550 BC).
The Hamangia culture is a Late Neolithic archaeological culture of Dobruja (Romania and Bulgaria) between the Danube and the Black Sea and Muntenia in the south.
It is named after the site of Baia-Hamangia, discovered in 1952 along Lake Golovita.[1]
http://upload.wikimedia.org/wikipedia/commons/f/f4/Hamangia_Muzeul_din_Constanta.JPG
http://upload.wikimedia.org/wikipedia/commons/1/10/European_Middle_Neolithic.gif
http://en.wikipedia.org/wiki/Durankulak
History[edit]
The Durankulak settlement commenced on a small island, approximately 5200 BC and lasted for thousand years.
The first inhabitants were the Hamangia culture, dated from the middle of the 6th millennium to the middle of 5th millennium BC,
and were the first manifestation of the Neolithic life in Dobruzha.
Hamangia people were small-scale cultivators and plant collectors who built houses, made pottery, herded and hunted animals
Durankulak is the north-easternmost inhabited place in Bulgaria and the northernmost village of the Bulgarian Black Sea Coast,
although the village itself is slightly inland. Durankulak lies north of the town of Shabla,
with the only places to the north along the coast being the formerly exclusively Czechoslovak camping site Kosmos and the Kartalburun and Sivriburun headlands.
Durankulak is also the name of the nearby border checkpoint on the Bulgarian-Romanian border; just north of the border is the Romanian seaside resort Vama Veche.
http://en.wikipedia.org/wiki/Hamangia_culture
The Thinker of Hamangia, Neolithic Hamangia culture (c. 5250-4550 BC).
The Hamangia culture is a Late Neolithic archaeological culture of Dobruja (Romania and Bulgaria) between the Danube and the Black Sea and Muntenia in the south.
It is named after the site of Baia-Hamangia, discovered in 1952 along Lake Golovita.[1]
http://upload.wikimedia.org/wikipedia/commons/f/f4/Hamangia_Muzeul_din_Constanta.JPG
{kind=link}
http://upload.wikimedia.org/wikipedia/commons/1/10/European_Middle_Neolithic.gif
{kind=link}
>>725
http://upload.wikimedia.org/wikipedia/commons/4/4d/Lake-Varna-map-EN.gif
Chalcolithic
http://en.wikipedia.org/wiki/Chalcolithic
The Chalcolithic period or Copper Age,[1] also known as the Eneolithic[1]/Aneolithic (from Latin aeneus "of bronze"),
is a phase of the Bronze Age before metallurgists discovered that adding tin to copper formed the harder bronze.
The Copper Age was originally defined as a transition between the Neolithic and the Bronze Age.
However, because it is characterized by the use of metals, the Copper Age is considered a part of the Bronze Age rather than the Stone Age.
The archaeological site of Belovode on the Rudnik mountain in Serbia contains the world's oldest securely dated evidence of copper making at high temperature, from 5,000 BC.[2][3]
Europe
An archaeological site in southeastern Europe (Serbia) contains the oldest securely dated evidence of copper making at high temperature, from 7,500 years ago.
The find in June 2010 extends the known record of copper smelting by about 800 years,
and suggests that copper smelting may have been invented in separate parts of Asia and Europe at that time rather than spreading from a single source
http://upload.wikimedia.org/wikipedia/commons/1/1c/Los_Millares_recreacion_cuadro.jpg
Painting of a Copper Age walled city, Los Millares, Iberia
http://upload.wikimedia.org/wikipedia/commons/4/4d/Lake-Varna-map-EN.gif
{kind=link}
Chalcolithic
http://en.wikipedia.org/wiki/Chalcolithic
The Chalcolithic period or Copper Age,[1] also known as the Eneolithic[1]/Aneolithic (from Latin aeneus "of bronze"),
is a phase of the Bronze Age before metallurgists discovered that adding tin to copper formed the harder bronze.
The Copper Age was originally defined as a transition between the Neolithic and the Bronze Age.
However, because it is characterized by the use of metals, the Copper Age is considered a part of the Bronze Age rather than the Stone Age.
The archaeological site of Belovode on the Rudnik mountain in Serbia contains the world's oldest securely dated evidence of copper making at high temperature, from 5,000 BC.[2][3]
Europe
An archaeological site in southeastern Europe (Serbia) contains the oldest securely dated evidence of copper making at high temperature, from 7,500 years ago.
The find in June 2010 extends the known record of copper smelting by about 800 years,
and suggests that copper smelting may have been invented in separate parts of Asia and Europe at that time rather than spreading from a single source
http://upload.wikimedia.org/wikipedia/commons/1/1c/Los_Millares_recreacion_cuadro.jpg
{kind=link}
Painting of a Copper Age walled city, Los Millares, Iberia
>>722
鰯もじゃなかったっけ?
鰯もじゃなかったっけ?
{kind=link}
733 :世界@名無史さん:2014/01/17(金) 11:11:53.82 0
【イギリスの移民】 2030年までに内戦か
http://www.youtube.com/watch?v=HViVQaPLktA・・・動画You Tube 3分32秒
【フランスの移民】 移民亡国
http://www.youtube.com/watch?v=LaRUAA5nwX8・・・動画You Tube 9分46秒
http://www.youtube.com/watch?v=oN7XvYPYU_U・・・動画You Tube 5分42秒
【フィンランドの移民】
http://www.youtube.com/watch?v=ZGEaUWKxRSs・・・動画You Tube 2分30秒
【イタリアの移民】 チャイナタウンでの暴動
http://www.youtube.com/watch?v=XkFhqQFrAus・・・動画You Tube 4分38秒
http://www.youtube.com/watch?v=HViVQaPLktA・・・動画You Tube 3分32秒
【フランスの移民】 移民亡国
http://www.youtube.com/watch?v=LaRUAA5nwX8・・・動画You Tube 9分46秒
http://www.youtube.com/watch?v=oN7XvYPYU_U・・・動画You Tube 5分42秒
【フィンランドの移民】
http://www.youtube.com/watch?v=ZGEaUWKxRSs・・・動画You Tube 2分30秒
【イタリアの移民】 チャイナタウンでの暴動
http://www.youtube.com/watch?v=XkFhqQFrAus・・・動画You Tube 4分38秒
幕末・明治、来日した外国人は日本人のブサイクさに驚愕したという
http://galasoku.livedoor.biz/archives/2565912.html
http://galasoku.livedoor.biz/archives/2565912.html
チェヘラアバード岩塩鉱
Saltmen ソルトマン
http://en.wikipedia.org/wiki/Saltmen
The Saltmen were discovered in the Chehrabad salt mines, located on the southern part of the Hamzehlu village, on the west side of the city of Zanjan, in the Zanjan Province in Iran.
Saltmen ソルトマン
http://en.wikipedia.org/wiki/Saltmen
The Saltmen were discovered in the Chehrabad salt mines, located on the southern part of the Hamzehlu village, on the west side of the city of Zanjan, in the Zanjan Province in Iran.
737 :世界@名無史さん:2014/01/17(金) 22:19:32.30 0
まさにお塩先生
>>730
Plo?nik
http://en.wikipedia.org/wiki/Plo%C4%8Dnik
Not long ago, an important European-archaeology excavation site was found in Plo?nik.
At this site, and in Belovode, archaeologists have found the earliest current evidence of copper smelting, dating from between 5500 BCE and 5000 BCE.[1][2]
This shows that the Copper Age started 500 years earlier than previously thought, and probably somewhere near this region.
Plo?nik
http://en.wikipedia.org/wiki/Plo%C4%8Dnik
Not long ago, an important European-archaeology excavation site was found in Plo?nik.
At this site, and in Belovode, archaeologists have found the earliest current evidence of copper smelting, dating from between 5500 BCE and 5000 BCE.[1][2]
This shows that the Copper Age started 500 years earlier than previously thought, and probably somewhere near this region.
>>735
Vin?a culture
http://en.wikipedia.org/wiki/Vin%C4%8Da_culture
The Vin?a culture, also known as Turda? culture or Turda?-Vin?a culture, is a Neolithic archaeological culture in Southeastern Europe, dated to the period 5500?4500 BCE
Geography and Demographics[edit]
The Vin?a culture occupied a region of Southeastern Europe (i.e. the Balkans) corresponding mainly to modern-day Serbia,
but also parts of Romania, Bulgaria, Bosnia, Montenegro, Macedonia, and Greece.[1]
This region had already been settled by farming societies of the First Temperate Neolithic,
but during the Vin?a period sustained population growth led to an unprecedented level of settlement size and density along with the population of areas that were bypassed by earlier settlers.
Vin?a settlements were considerably larger than any other contemporary European culture,
in some instances surpassing the cities of the Aegean and early Near Eastern Bronze Age a millennium later.
The largest sites?some more than 300,000 square metres?may have been home to up to 2,500 people.[2]
http://upload.wikimedia.org/wikipedia/commons/6/60/Vin%C4%8Da_culture_locator_map.svg
http://upload.wikimedia.org/wikipedia/commons/0/00/Vincha_066.jpg
http://upload.wikimedia.org/wikipedia/commons/5/5b/Lady_of_Vinca.jpg
Neolithic clay amulet (retouched), part of the T?rt?ria tablets set, dated to 5500-5300 BC and associated with the Turda?-Vin?a culture.
The Vin?a symbols on it predate the proto-Sumerian pictographic script. Discovered in 1961 at T?rt?ria by the archaeologist Nicolae Vlassa.
http://upload.wikimedia.org/wikipedia/commons/4/4f/Tartaria_amulet_retouched.PNG
Vin?a culture
http://en.wikipedia.org/wiki/Vin%C4%8Da_culture
The Vin?a culture, also known as Turda? culture or Turda?-Vin?a culture, is a Neolithic archaeological culture in Southeastern Europe, dated to the period 5500?4500 BCE
Geography and Demographics[edit]
The Vin?a culture occupied a region of Southeastern Europe (i.e. the Balkans) corresponding mainly to modern-day Serbia,
but also parts of Romania, Bulgaria, Bosnia, Montenegro, Macedonia, and Greece.[1]
This region had already been settled by farming societies of the First Temperate Neolithic,
but during the Vin?a period sustained population growth led to an unprecedented level of settlement size and density along with the population of areas that were bypassed by earlier settlers.
Vin?a settlements were considerably larger than any other contemporary European culture,
in some instances surpassing the cities of the Aegean and early Near Eastern Bronze Age a millennium later.
The largest sites?some more than 300,000 square metres?may have been home to up to 2,500 people.[2]
http://upload.wikimedia.org/wikipedia/commons/6/60/Vin%C4%8Da_culture_locator_map.svg
{kind=link}
http://upload.wikimedia.org/wikipedia/commons/0/00/Vincha_066.jpg
{kind=link}
http://upload.wikimedia.org/wikipedia/commons/5/5b/Lady_of_Vinca.jpg
{kind=link}
Neolithic clay amulet (retouched), part of the T?rt?ria tablets set, dated to 5500-5300 BC and associated with the Turda?-Vin?a culture.
The Vin?a symbols on it predate the proto-Sumerian pictographic script. Discovered in 1961 at T?rt?ria by the archaeologist Nicolae Vlassa.
http://upload.wikimedia.org/wikipedia/commons/4/4f/Tartaria_amulet_retouched.PNG
{kind=link}
740 :世界@名無史さん:2014/01/18(土) 08:53:23.33 0
>>735
遺伝子サイトみると、これがR1bらしい。
遺伝子サイトみると、これがR1bらしい。
741 :世界@名無史さん:2014/01/18(土) 10:48:41.71 0
バルカンのI2が無傷だったことを考えると、ドナウ川を遡って、あまり脇道にはそれなかったってことか
フィクションで時代も違うけど、氷河期が舞台の「エイラ-地上の旅人」シリーズで、ジョンダラーとソノーランの兄弟が辿ったルートもこの道だな。逆方向だけど
大昔から重要なルートか
フィクションで時代も違うけど、氷河期が舞台の「エイラ-地上の旅人」シリーズで、ジョンダラーとソノーランの兄弟が辿ったルートもこの道だな。逆方向だけど
大昔から重要なルートか
742 :世界@名無史さん:2014/01/18(土) 11:03:11.09 0
【スイス】 外国人犯罪者の国外追放を義務化する国民投票
http://www.youtube.com/watch?v=Y7FWWJwLMGg・・・動画You Tube/3分21秒
【スペイン】 移民により闘牛が違法になった都市
http://www.youtube.com/watch?v=6U6qaUgodzk・・・動画You Tube/4分56秒
【ベルギー】 移民の人口が25%以上になった首都
http://www.youtube.com/watch?v=VdHEAG8DEP4・・・動画You Tube/5分40秒
http://www.youtube.com/watch?v=Y7FWWJwLMGg・・・動画You Tube/3分21秒
【スペイン】 移民により闘牛が違法になった都市
http://www.youtube.com/watch?v=6U6qaUgodzk・・・動画You Tube/4分56秒
【ベルギー】 移民の人口が25%以上になった首都
http://www.youtube.com/watch?v=VdHEAG8DEP4・・・動画You Tube/5分40秒
The Bronze Age Indo-European invasion of Europe
http://dienekes.blogspot.com.es/2012/07/bronze-age-indo-european-invasion-of.html
We can plainly see that European hunter-gatherers best map to the modern Atlantic_Baltic population component.
This is well represented in the remotest areas of Europe, the ones most distant from the Near Eastern womb of nations.
It can be reasonably supposed that the modern Atlantic_Baltic component partially captures alleles present in the ancient European hunter-gatherers,
the mtDNA haplogroup U population that seems to stretch from Iberia to Siberia.
(However, note, that this does not mean that the Atlantic_Baltic component represents hunter-gatherer ancestry only.)
http://4.bp.blogspot.com/-ZwmxlhXO8-k/T_B9lrlnFJI/AAAAAAAAE7w/My33JaMKr_E/s1600/ancientdna12.png
'K12b' and 'K7b' calculators
http://dodecad.blogspot.com.es/2012/01/k12b-and-k7b-calculators.html
なはっは、発掘サイトの名前じゃなかったww
http://dienekes.blogspot.com.es/2012/07/bronze-age-indo-european-invasion-of.html
We can plainly see that European hunter-gatherers best map to the modern Atlantic_Baltic population component.
This is well represented in the remotest areas of Europe, the ones most distant from the Near Eastern womb of nations.
It can be reasonably supposed that the modern Atlantic_Baltic component partially captures alleles present in the ancient European hunter-gatherers,
the mtDNA haplogroup U population that seems to stretch from Iberia to Siberia.
(However, note, that this does not mean that the Atlantic_Baltic component represents hunter-gatherer ancestry only.)
http://4.bp.blogspot.com/-ZwmxlhXO8-k/T_B9lrlnFJI/AAAAAAAAE7w/My33JaMKr_E/s1600/ancientdna12.png
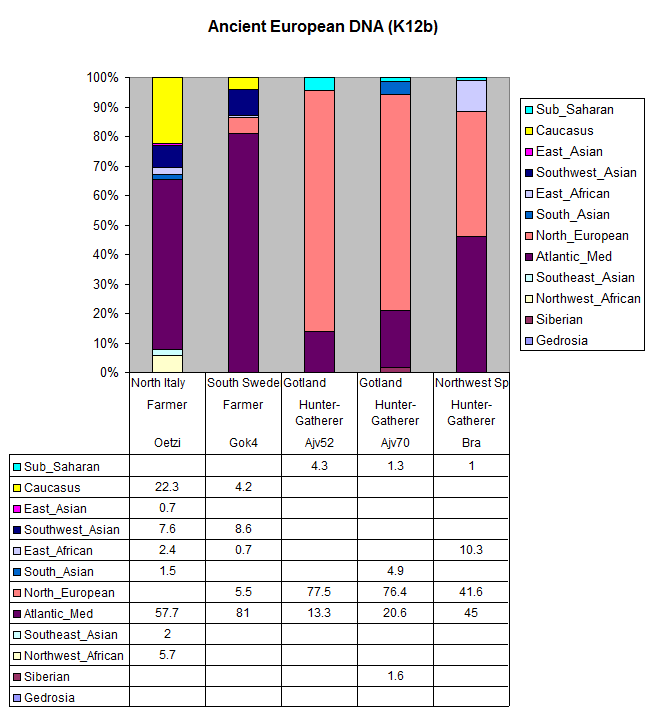{kind=link}
'K12b' and 'K7b' calculators
http://dodecad.blogspot.com.es/2012/01/k12b-and-k7b-calculators.html
なはっは、発掘サイトの名前じゃなかったww
>>548>>574
Ancient mtDNA maps of Europe
http://forwhattheywereweare.blogspot.jp/p/ancient-mtdna-maps-of-europe.html
これは非常に良い気がする
ただ、Hが良くわからない
Ancient mtDNA maps of Europe
http://forwhattheywereweare.blogspot.jp/p/ancient-mtdna-maps-of-europe.html
これは非常に良い気がする
ただ、Hが良くわからない
>>740>>741
外人も含めてみんな、
Yのハプロばっかり好きなんだよなww
Ancient European DNA and some debatable conclusions (からHaploid DNA抜粋)
The new European hunter-gatherer samples carried all Y-DNA I and mtDNA U5a and U2e.
More specifically, the hunter-gatherer mtDNA lineages are:
?Lochsbour (Luxembourg): U5b1a
?Motala (Sweden):
?Motala 1 & 3: U5b1a
?Motala 2 & 12: U2e1
?Motala 4 & 6: U5a2d
?Motala 9: U5a2
Additionally the Stuttgart Linear Pottery farmer (female) carried the mtDNA lineage T2c1d1.
The Y-DNA lineages are:
?Lochsbour: I2a1b*(xI2a1b1, I2a1b2, I2a1b3)
?Motala 2: I*(xI1, I2a2,I2a1b3)
?Motala 3: I2*(xI2a1a, I2a2, I2b)
?Motala 6: uncertain (L55+ would make it Q1a2a but L232- forces it out of Q1)
?Motala 9: I*(xI1)
?Motala 12: I2a1b*(xI2a1b1, I2a1b3)
These are with certainty the oldest Y-DNA sequences of Europe so far and the fact that all them fall within haplogroup I(xI1) supports the notion of this lineage being once common in the subcontinent, at least in some areas.
Today I2 is most common in Sardinia, the NW Balcans (Croatia, Bosnia, Montenegro), North Germany and areas around Moldavia.
http://4.bp.blogspot.com/-ufx6McTzLmA/UrwiBjBR8II/AAAAAAAACVA/uY4XqHvhNzQ/s1600/PCAannot4.png
外人も含めてみんな、
Yのハプロばっかり好きなんだよなww
Ancient European DNA and some debatable conclusions (からHaploid DNA抜粋)
The new European hunter-gatherer samples carried all Y-DNA I and mtDNA U5a and U2e.
More specifically, the hunter-gatherer mtDNA lineages are:
?Lochsbour (Luxembourg): U5b1a
?Motala (Sweden):
?Motala 1 & 3: U5b1a
?Motala 2 & 12: U2e1
?Motala 4 & 6: U5a2d
?Motala 9: U5a2
Additionally the Stuttgart Linear Pottery farmer (female) carried the mtDNA lineage T2c1d1.
The Y-DNA lineages are:
?Lochsbour: I2a1b*(xI2a1b1, I2a1b2, I2a1b3)
?Motala 2: I*(xI1, I2a2,I2a1b3)
?Motala 3: I2*(xI2a1a, I2a2, I2b)
?Motala 6: uncertain (L55+ would make it Q1a2a but L232- forces it out of Q1)
?Motala 9: I*(xI1)
?Motala 12: I2a1b*(xI2a1b1, I2a1b3)
These are with certainty the oldest Y-DNA sequences of Europe so far and the fact that all them fall within haplogroup I(xI1) supports the notion of this lineage being once common in the subcontinent, at least in some areas.
Today I2 is most common in Sardinia, the NW Balcans (Croatia, Bosnia, Montenegro), North Germany and areas around Moldavia.
http://4.bp.blogspot.com/-ufx6McTzLmA/UrwiBjBR8II/AAAAAAAACVA/uY4XqHvhNzQ/s1600/PCAannot4.png
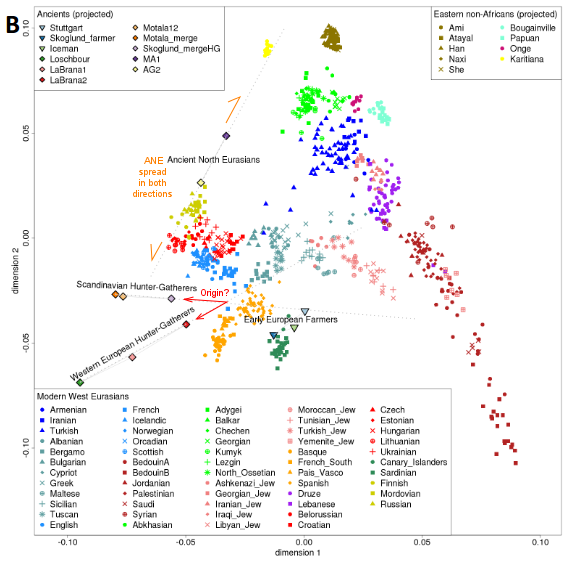{kind=link}
747 :世界@名無史さん:2014/01/18(土) 13:15:27.38 0
>>735
アイスマンの持っていた銅斧の産地はどこか?
http://www3.kmu.ac.jp/legalmed/DNA/iceman.html
名称はエッツィにしたいがめんどくさそうなので
ちなみに、へぼい検索能力ではこれしか見つからなかった
謎を掛けるオーストリアのミイラ
http://lang-8.com/97862/journals/1157498
肩に刺さる矢が死亡の原因である可能性が高いという。この2.5センチの石鏃はCTスキャンで念入りに調べ直し、原料と原産地を知ることかできた。
研究者によると、原料がヴェローナの北にあるモンディレシーニからのものだ。表面処理の方法によって、 投射物が北イタリアのレモデッロ文化に所属できる。
但し、そのような有茎式(ゆうくきしき)の鏃北アルプスにあるモンド湖でも流通していたので、加害者がどこから生まれてきたのだろうかまだ分からないものだ。
エッツィが持っていた銅斧はレモデッロ文化にも所属できるが、原料地はオーストリアのモンド湖です。
そのためアイスマンは石器時代の銅探検隊の首領かもしれない。
銅を持ち帰った時には、追剥に襲われ、エッツィは致命傷を負って、現地で埋葬した。
アイスマンの持っていた銅斧の産地はどこか?
http://www3.kmu.ac.jp/legalmed/DNA/iceman.html
名称はエッツィにしたいがめんどくさそうなので
ちなみに、へぼい検索能力ではこれしか見つからなかった
謎を掛けるオーストリアのミイラ
http://lang-8.com/97862/journals/1157498
肩に刺さる矢が死亡の原因である可能性が高いという。この2.5センチの石鏃はCTスキャンで念入りに調べ直し、原料と原産地を知ることかできた。
研究者によると、原料がヴェローナの北にあるモンディレシーニからのものだ。表面処理の方法によって、 投射物が北イタリアのレモデッロ文化に所属できる。
但し、そのような有茎式(ゆうくきしき)の鏃北アルプスにあるモンド湖でも流通していたので、加害者がどこから生まれてきたのだろうかまだ分からないものだ。
エッツィが持っていた銅斧はレモデッロ文化にも所属できるが、原料地はオーストリアのモンド湖です。
そのためアイスマンは石器時代の銅探検隊の首領かもしれない。
銅を持ち帰った時には、追剥に襲われ、エッツィは致命傷を負って、現地で埋葬した。
>>682>>635
Prehistory of Corsica
http://en.wikipedia.org/wiki/Prehistory_of_Corsica
俺の中ではコルシカなんだが、、、
The insular prehistory of Corsica begins with the Mesolithic (Pre-Neolithic)
when people from prehistoric Sardinia crossed the Strait of Bonifacio to hunt from rock shelters in Corsica at approximately 9000 BC.
It ends with colonization by the Ancient Greeks at Aleria in 566 BC,
the Iron Age. Corsica, or Kyrnos, is not mentioned before then. Thus the history of Corsica begins in 566 BC.
Genetic demographics
For Corte, which is inland Corsica, the investigators use the term stochastic (random),
finding a higher variability among Corsicans than within the rest of the Mediterranean reference population.
There was also a gap between Corse-du-Sud and Haute-Corse.
Using an assumed generation of 25 years, the investigators estimate a base population at 20,000 BP[clarification needed] for the first humans of Corsica.
As this number is in no way supported by archaeological evidence, it suggests that cultural remains of the Paleolithic may well yet turn up.
> The greatest genetic distances were between the Sardinian and Corsican populations, which indicates that the islands were settled by different people.
The investigators exclude any significant gene flow between the two islands.
> The closest affinities were with the population of Tuscany.
The investigators attribute this closer kinship to a major influx from there in the early and middle Neolithic, which they date to the 8th-6th millennia BP.
The separation of Corsican and non-Corsican populations falls within rather wide limits: no earlier than 19,000 BP or later than between 4929 and 8746 BP.
An indigenous Corsican population had therefore formed by about 3000 BC at latest, in this view.
Prehistory of Corsica
http://en.wikipedia.org/wiki/Prehistory_of_Corsica
俺の中ではコルシカなんだが、、、
The insular prehistory of Corsica begins with the Mesolithic (Pre-Neolithic)
when people from prehistoric Sardinia crossed the Strait of Bonifacio to hunt from rock shelters in Corsica at approximately 9000 BC.
It ends with colonization by the Ancient Greeks at Aleria in 566 BC,
the Iron Age. Corsica, or Kyrnos, is not mentioned before then. Thus the history of Corsica begins in 566 BC.
Genetic demographics
For Corte, which is inland Corsica, the investigators use the term stochastic (random),
finding a higher variability among Corsicans than within the rest of the Mediterranean reference population.
There was also a gap between Corse-du-Sud and Haute-Corse.
Using an assumed generation of 25 years, the investigators estimate a base population at 20,000 BP[clarification needed] for the first humans of Corsica.
As this number is in no way supported by archaeological evidence, it suggests that cultural remains of the Paleolithic may well yet turn up.
> The greatest genetic distances were between the Sardinian and Corsican populations, which indicates that the islands were settled by different people.
The investigators exclude any significant gene flow between the two islands.
> The closest affinities were with the population of Tuscany.
The investigators attribute this closer kinship to a major influx from there in the early and middle Neolithic, which they date to the 8th-6th millennia BP.
The separation of Corsican and non-Corsican populations falls within rather wide limits: no earlier than 19,000 BP or later than between 4929 and 8746 BP.
An indigenous Corsican population had therefore formed by about 3000 BC at latest, in this view.
>>691
エトルリア
http://en.wikipedia.org/wiki/Etruscan_civilization
The newest mtDNA study confirms that the Etruscans were not related substantially to the Upper Paleolithic hunter-gatherer populations of Europe,
and also showed no similarities to populations in the Near East. Another earlier DNA study performed in Italy however, partly gave credence to the theory of Herodotus,
as the results showed 11 minor mitochondrial DNA lineages extracted from different Etruscan remains occur nowhere else in Europe, and are shared only with Near East people.[9]
共同墓地は、実際に古代に定住があったことを証明している。
海の近く、海岸から数キロメートルのところに集落を形成する一般の傾向に反して、隔離された、住むのに適さない、変わった内陸の環境に居を定めているように見て取れる。
唯一のエトルリア海洋都市は、おそらくポプロニアであり、ほかは後背地に位置をとっているが、これは海賊の来襲を恐れたためである。
すなわち、クレタ人やミケーネ人に代わって、何者かが鉄を求めてこの資源豊かなエトルリアの土地、イタリア沿岸を訪れていたことがわかる。
とにかく、ポプロニアのように海に面した、それもエルバ島の正面という場所柄には理由がある。
ここはおそらく、ヴィッラノーヴァ期の銅・銀製品の主要積出港であり、後にエトルリア期に入って、“鉄の港”となったのである。
研究者たちが偽アリストテレスと呼ぶ 古代の無名著作家は、ポプロニアでは銅を産出したと記しており、実際に銅くずや、加工工場の跡が見られる。
のちにポプロニアは、エルバ島産の鉄の加工で非常に重要な地となった。
エトルリア
http://en.wikipedia.org/wiki/Etruscan_civilization
The newest mtDNA study confirms that the Etruscans were not related substantially to the Upper Paleolithic hunter-gatherer populations of Europe,
and also showed no similarities to populations in the Near East. Another earlier DNA study performed in Italy however, partly gave credence to the theory of Herodotus,
as the results showed 11 minor mitochondrial DNA lineages extracted from different Etruscan remains occur nowhere else in Europe, and are shared only with Near East people.[9]
共同墓地は、実際に古代に定住があったことを証明している。
海の近く、海岸から数キロメートルのところに集落を形成する一般の傾向に反して、隔離された、住むのに適さない、変わった内陸の環境に居を定めているように見て取れる。
唯一のエトルリア海洋都市は、おそらくポプロニアであり、ほかは後背地に位置をとっているが、これは海賊の来襲を恐れたためである。
すなわち、クレタ人やミケーネ人に代わって、何者かが鉄を求めてこの資源豊かなエトルリアの土地、イタリア沿岸を訪れていたことがわかる。
とにかく、ポプロニアのように海に面した、それもエルバ島の正面という場所柄には理由がある。
ここはおそらく、ヴィッラノーヴァ期の銅・銀製品の主要積出港であり、後にエトルリア期に入って、“鉄の港”となったのである。
研究者たちが偽アリストテレスと呼ぶ 古代の無名著作家は、ポプロニアでは銅を産出したと記しており、実際に銅くずや、加工工場の跡が見られる。
のちにポプロニアは、エルバ島産の鉄の加工で非常に重要な地となった。
>>741
確かにドナウに見える、、、
大昔の採集民と農耕民はうまくやっていた
http://yuihaga.blog.fc2.com/blog-entry-243.html
文化交流の可能性に集中するため、カーディフ大学(英国)の考古学者、ドゥシャン・ボリッチ(Du?an Bori?)とウィスコンシン大学(マディソン)の考古学者、T・ダグラス・プライスは
ドナウ峡谷にある9遺跡から産した153体の骨格の、歯におけるストロンチウム同位体の比率を分析した。
これらの骨格は13,500から7500年前、すなわち中石器時代のすぐ前から新石器時代初期まで、と年代測定されている。
ストロンチウム87とストロンチウム86の同位体比率は異なった土壌と岩石で大きく変化し、摂取されたストロンチウムは子供の歯に集積され、
ある人物がどこで成長したかについての一生涯のシグネチャを提供する。
歯と地元の環境のストロンチウム比率を比較することによって、研究者たちは誰がある特定の地域で成長した人で、誰が別の場所で成長した移住者かを見分けられる。
ボリッチとプライスは直接炭素年代測定によって年代が直接決定されてる骨格を分析し、
新石器文化の最初のサインが見られる頃にストロンチウム組成に大きな変化があったことを突き止めた。
彼らは今週それをProceedings of the National Academy of Sciences誌に報告した。
ドナウ川沿いの中石器人のストロンチウム同位体比は彼らが生活をしていた川の堆積物を反映したものだった。
しかし8200年前、これより著しく高い比率と低い比率をもつ骨格が出現する。
これらの骨格は文化が元来の中石器のままに見える遺跡にすら現れる。「非地元」な個体はドナウ川から遠く離れた地域から来たように思われ、移住者の数は時間を経るとともにジャンプする。
たとえば8200年前より前は、レペンスキ・ヴィールのほぼすべての埋葬者が地元民だった。
しかし8200年から7950年前までの間に埋葬された19人のうち5人は非地元民だった――そしてすべてが女性だった。
確かにドナウに見える、、、
大昔の採集民と農耕民はうまくやっていた
http://yuihaga.blog.fc2.com/blog-entry-243.html
文化交流の可能性に集中するため、カーディフ大学(英国)の考古学者、ドゥシャン・ボリッチ(Du?an Bori?)とウィスコンシン大学(マディソン)の考古学者、T・ダグラス・プライスは
ドナウ峡谷にある9遺跡から産した153体の骨格の、歯におけるストロンチウム同位体の比率を分析した。
これらの骨格は13,500から7500年前、すなわち中石器時代のすぐ前から新石器時代初期まで、と年代測定されている。
ストロンチウム87とストロンチウム86の同位体比率は異なった土壌と岩石で大きく変化し、摂取されたストロンチウムは子供の歯に集積され、
ある人物がどこで成長したかについての一生涯のシグネチャを提供する。
歯と地元の環境のストロンチウム比率を比較することによって、研究者たちは誰がある特定の地域で成長した人で、誰が別の場所で成長した移住者かを見分けられる。
ボリッチとプライスは直接炭素年代測定によって年代が直接決定されてる骨格を分析し、
新石器文化の最初のサインが見られる頃にストロンチウム組成に大きな変化があったことを突き止めた。
彼らは今週それをProceedings of the National Academy of Sciences誌に報告した。
ドナウ川沿いの中石器人のストロンチウム同位体比は彼らが生活をしていた川の堆積物を反映したものだった。
しかし8200年前、これより著しく高い比率と低い比率をもつ骨格が出現する。
これらの骨格は文化が元来の中石器のままに見える遺跡にすら現れる。「非地元」な個体はドナウ川から遠く離れた地域から来たように思われ、移住者の数は時間を経るとともにジャンプする。
たとえば8200年前より前は、レペンスキ・ヴィールのほぼすべての埋葬者が地元民だった。
しかし8200年から7950年前までの間に埋葬された19人のうち5人は非地元民だった――そしてすべてが女性だった。
>>754
ボリッチとプライスは非地元民は中石器文化の移住者のものであると結論づける。
彼らの、人物の食性を表す、歯と骨における窒素同位体比率はその考えを裏打ちする。
非地元民は海産物より陸産物の食性に典型的な窒素比率を持つ傾向があった。これは魚を食べない農耕民に予想されることだ。
埋葬者に中石器人と新石器人が混じり合ってることは、新石器文化が中石器文化を置き換えるまでの少なくとも200年にわたって、
2つのグループが隣り合って生活していたことを示す、とボリッチとプライスは結論する。
「採集民コミュニティは新石器社会ネットワークと敵対するのではなく、その中に組み込まれていった」とボリッチは話す。
「この図は大部分において平和な共存の一つだ」
ブリストル大学(英国)の考古学者、アレクス・ベントリー(Alex Bentley)はこの研究は「偉大な論文」だと呼ぶ。
彼は新石器人の女性が中石器コミュニティへと移住していったことを示す証拠は「すごく面白い」と続ける。
以前の遺伝的研究は農耕が西に向かってヨーロッパへと広がるとともに反対のことが起こったことを示していたからだ
――すなわち、狩猟採集民の女の子が新石器コミュニティに入って農耕民の男の子と結婚したということ。
実際にユニヴァーシティ・カレッジ・ダブリンの考古学者、ロン・ピンハシ(Ron Pinhasi)は、
このドナウ峡谷のシナリオは中石器-新石器移行期のヨーロッパ全域のモデルにはならないだろう、と注意する。
「私はこれらの集団を[採集民-農耕民交流の典型的な例ではなく]孤立例だと見ている」とピンハシは話す。
Science NOW
Ancient Foragers and Farmers Hit It Off
Michael Balter, 11 February 2013
原論文
Du?an Bori? and T. Douglas Price
Strontium isotopes document greater human mobility at the start of the Balkan Neolithic
PNAS February 11, 2013, Published online before print February 11, 2013
doi: 10.1073/pnas.1211474110
ボリッチとプライスは非地元民は中石器文化の移住者のものであると結論づける。
彼らの、人物の食性を表す、歯と骨における窒素同位体比率はその考えを裏打ちする。
非地元民は海産物より陸産物の食性に典型的な窒素比率を持つ傾向があった。これは魚を食べない農耕民に予想されることだ。
埋葬者に中石器人と新石器人が混じり合ってることは、新石器文化が中石器文化を置き換えるまでの少なくとも200年にわたって、
2つのグループが隣り合って生活していたことを示す、とボリッチとプライスは結論する。
「採集民コミュニティは新石器社会ネットワークと敵対するのではなく、その中に組み込まれていった」とボリッチは話す。
「この図は大部分において平和な共存の一つだ」
ブリストル大学(英国)の考古学者、アレクス・ベントリー(Alex Bentley)はこの研究は「偉大な論文」だと呼ぶ。
彼は新石器人の女性が中石器コミュニティへと移住していったことを示す証拠は「すごく面白い」と続ける。
以前の遺伝的研究は農耕が西に向かってヨーロッパへと広がるとともに反対のことが起こったことを示していたからだ
――すなわち、狩猟採集民の女の子が新石器コミュニティに入って農耕民の男の子と結婚したということ。
実際にユニヴァーシティ・カレッジ・ダブリンの考古学者、ロン・ピンハシ(Ron Pinhasi)は、
このドナウ峡谷のシナリオは中石器-新石器移行期のヨーロッパ全域のモデルにはならないだろう、と注意する。
「私はこれらの集団を[採集民-農耕民交流の典型的な例ではなく]孤立例だと見ている」とピンハシは話す。
Science NOW
Ancient Foragers and Farmers Hit It Off
Michael Balter, 11 February 2013
原論文
Du?an Bori? and T. Douglas Price
Strontium isotopes document greater human mobility at the start of the Balkan Neolithic
PNAS February 11, 2013, Published online before print February 11, 2013
doi: 10.1073/pnas.1211474110
ヨーロッパ最初の農耕民は移住者だった(抜粋)
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=93566949
ヨーロッパの中央部や西部で農耕を始めたのは、やりから農具へと徐々に道具を持ち替えていった土着の狩猟採集民族ではなく、
紀元前5500年ごろに家畜と一緒に移住してきた外部の農耕民族だったという。しかし、先住民はよそ者たちをどうやら歓迎しなかったらしい。
ドイツやリトアニア、ポーランド、ロシアで発掘された古代の狩猟採集民族と初期農耕民族の古人骨から遺伝物質を採取し分析した結果、このような結論が導き出されたという。
農耕文化は約7500年前に急拡大し、西はフランス西部あたりにまで伝わったと一般には考えられている。
今回の研究成果は、最後の氷河期が終わった約1万年前にヨーロッパに定住した狩猟採集民族が同地域最初の農耕民族に転身したとする昔からの通説と相容れないものだ。
ブルガー氏の研究チームは、狩猟採集民族と初期農耕民族は当時まだ隔絶した状態で生活していたことを突き止め、9月4日発行の「Science」誌で発表した。
「両者の間に“文化”の交流はあっただろうが、遺伝子分析の結果を見る限り、少なくとも当初は“血”の交流はなかったと思われる。
彼らは異なる人種で、狩猟採集民族と農耕民族という2つの社会が並立していたということだ」と同氏は解説する。
研究チームは両者の古人骨を次のように区別した。
8000年以上前、つまりヨーロッパで農耕が始まる前のものや、矢じり、クマの歯の首飾りといった狩猟に関係する遺物が周囲で見つかったものは狩猟採集民族に分類し、
農耕具や家畜の骨と一緒に発見されたものは農耕民族に分類したという。
そしてそれらの古人骨から採取した遺伝物質と現代ヨーロッパ人の遺伝子を比較したところ、ある謎が浮かび上がった。
2つの血統だけでは、現代ヨーロッパ人の遺伝子プールは完成しないのである。
その結果、現代ヨーロッパ人の起源がほとんど解明されていない現状が浮き彫りになった。
「別の人種の移住か、遺伝的変異か。とにかく未発見の要素が何かしらあるはずだ」とブルガー氏は言う。
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=93566949
ヨーロッパの中央部や西部で農耕を始めたのは、やりから農具へと徐々に道具を持ち替えていった土着の狩猟採集民族ではなく、
紀元前5500年ごろに家畜と一緒に移住してきた外部の農耕民族だったという。しかし、先住民はよそ者たちをどうやら歓迎しなかったらしい。
ドイツやリトアニア、ポーランド、ロシアで発掘された古代の狩猟採集民族と初期農耕民族の古人骨から遺伝物質を採取し分析した結果、このような結論が導き出されたという。
農耕文化は約7500年前に急拡大し、西はフランス西部あたりにまで伝わったと一般には考えられている。
今回の研究成果は、最後の氷河期が終わった約1万年前にヨーロッパに定住した狩猟採集民族が同地域最初の農耕民族に転身したとする昔からの通説と相容れないものだ。
ブルガー氏の研究チームは、狩猟採集民族と初期農耕民族は当時まだ隔絶した状態で生活していたことを突き止め、9月4日発行の「Science」誌で発表した。
「両者の間に“文化”の交流はあっただろうが、遺伝子分析の結果を見る限り、少なくとも当初は“血”の交流はなかったと思われる。
彼らは異なる人種で、狩猟採集民族と農耕民族という2つの社会が並立していたということだ」と同氏は解説する。
研究チームは両者の古人骨を次のように区別した。
8000年以上前、つまりヨーロッパで農耕が始まる前のものや、矢じり、クマの歯の首飾りといった狩猟に関係する遺物が周囲で見つかったものは狩猟採集民族に分類し、
農耕具や家畜の骨と一緒に発見されたものは農耕民族に分類したという。
そしてそれらの古人骨から採取した遺伝物質と現代ヨーロッパ人の遺伝子を比較したところ、ある謎が浮かび上がった。
2つの血統だけでは、現代ヨーロッパ人の遺伝子プールは完成しないのである。
その結果、現代ヨーロッパ人の起源がほとんど解明されていない現状が浮き彫りになった。
「別の人種の移住か、遺伝的変異か。とにかく未発見の要素が何かしらあるはずだ」とブルガー氏は言う。
チーズづくりの技は7,500年の歴史がある(抜粋)
Nidhi Subbaraman
12 December 2012
http://yuihaga.blog.fc2.com/blog-entry-199.html
プリンストン大学(ニュージャージー州)の考古学者、ピーター・ボグスキ(Peter Bogucki)は1980年代に初めて
チーズ作りがヨーロッパで紀元前5,500に行われていたのではないかと考えた人の一人だった。
彼は現在はポーランドとなっているところの古代のウシ飼育遺構を研究する考古学者たちが、
チーズ漉し器を思わせる、穴だらけの土器の欠片を見つけていたことに気づいた。
ボグスキは新石器時代の農家が家畜を乳や肉以外に利用する方法を見つけていたと推論した。
乳製品文化
チーズ作りは新石器時代の農家に家畜から利用できる資源を最大限活用する方法を与えたと思われる。
初期のヒトは幼時を過ぎると乳糖(ラクトース)を消化することができなかった。
だが、伝統的なチーズには生乳よりはるかに少ない乳糖しか含まれない。
「チーズ作りは病気にならずに乳の不消化性を乗り越えられるようにしただろう」とエヴァシェドは話す。
Nidhi Subbaraman
12 December 2012
http://yuihaga.blog.fc2.com/blog-entry-199.html
プリンストン大学(ニュージャージー州)の考古学者、ピーター・ボグスキ(Peter Bogucki)は1980年代に初めて
チーズ作りがヨーロッパで紀元前5,500に行われていたのではないかと考えた人の一人だった。
彼は現在はポーランドとなっているところの古代のウシ飼育遺構を研究する考古学者たちが、
チーズ漉し器を思わせる、穴だらけの土器の欠片を見つけていたことに気づいた。
ボグスキは新石器時代の農家が家畜を乳や肉以外に利用する方法を見つけていたと推論した。
乳製品文化
チーズ作りは新石器時代の農家に家畜から利用できる資源を最大限活用する方法を与えたと思われる。
初期のヒトは幼時を過ぎると乳糖(ラクトース)を消化することができなかった。
だが、伝統的なチーズには生乳よりはるかに少ない乳糖しか含まれない。
「チーズ作りは病気にならずに乳の不消化性を乗り越えられるようにしただろう」とエヴァシェドは話す。
759 :世界@名無史さん:2014/01/20(月) 10:25:01.96 0
動画 You Tube 【ベルギー】最大のイスラム人口を抱える欧州の首都/アメリカのTV局作成
2分57秒・・・http://www.youtube.com/watch?v=td4SZ2Ao-9M
動画 You Tube 【イギリス】イスラム法の広がりをとめられないと宣言した移民
3分46秒・・・http://www.youtube.com/watch?v=ZRnyzxfGZa4
動画 You Tube 【フランス】フランスは20年後に旧植民地の植民地になる by プ−チン大統領
6分30秒・・・http://www.youtube.com/watch?v=qF8vgUFFN-g
2分57秒・・・http://www.youtube.com/watch?v=td4SZ2Ao-9M
動画 You Tube 【イギリス】イスラム法の広がりをとめられないと宣言した移民
3分46秒・・・http://www.youtube.com/watch?v=ZRnyzxfGZa4
動画 You Tube 【フランス】フランスは20年後に旧植民地の植民地になる by プ−チン大統領
6分30秒・・・http://www.youtube.com/watch?v=qF8vgUFFN-g
760 :世界@名無史さん:2014/01/20(月) 11:21:19.79 0
>>757 その後恒常的に生乳を飲まないといけない状況が起きたのはなんでだろう。乳を沸かして殺菌する燃料となる木材の枯渇?
761 :世界@名無史さん:2014/01/20(月) 19:16:17.60 0
とにかくR1bが欧州に侵入したのは4500〜5000年前だからね。
それまでに狩猟採集民と農耕民の最低ふたつの人類の流れがあるわけだ。
最後に大規模侵入したのが牧畜民R1a・R1b。
主要な侵入経路は少し前に書込みがあったドナウ北上ルート。
それまでに狩猟採集民と農耕民の最低ふたつの人類の流れがあるわけだ。
最後に大規模侵入したのが牧畜民R1a・R1b。
主要な侵入経路は少し前に書込みがあったドナウ北上ルート。
762 :世界@名無史さん:2014/01/20(月) 19:37:37.71 0
狩猟民の末裔と思われるIが北海周辺とバルカンに今もまとまって見られるのに対し、初期農耕民のマーカーと思われるY-DNAハプログループが顕著に見られないのはなんでだろう。
ジェファーソン一族ののTとかはそれっぽいけど
ジェファーソン一族ののTとかはそれっぽいけど
763 :世界@名無史さん:2014/01/20(月) 23:08:56.42 0
57 :名無しさんの主張:2014/01/20(月) 00:58:46.33 ID:/JcFFsuP
>>54
あと国民性がシャイでネクラだから西洋人の真似事=嫉妬と嘲笑の
対象にまだなっている。
ハングルやってる人間を僻まないでしょう??
眼中にもないだろうし。
63 :名無しさんの主張:2014/01/20(月) 04:36:28.68 ID:???
>>57
日本人同士で差別蔑視とか
お前は日本人だから腹立つとかムカつくわとか
訳の分からない事を平気で言い出す阿呆の陽気さとかは絡み辛いだけや(笑)
それとバカッターを筆頭に
陽気であれば何でも出来る(キリッって国民性なのが日本人(笑)だからなw
つまらない芸人に法的措置を取るよと言われて
社会主義だ全体主義だと叫ぶ馬鹿日本人(笑)
日本って反日本主義や反日本人主義なだけだと思うw社会主義にすらなってない。
日本や日本人が嫌いなのは分かるが
標準(笑)と変わらないのが大半だろうな。
>>54
あと国民性がシャイでネクラだから西洋人の真似事=嫉妬と嘲笑の
対象にまだなっている。
ハングルやってる人間を僻まないでしょう??
眼中にもないだろうし。
63 :名無しさんの主張:2014/01/20(月) 04:36:28.68 ID:???
>>57
日本人同士で差別蔑視とか
お前は日本人だから腹立つとかムカつくわとか
訳の分からない事を平気で言い出す阿呆の陽気さとかは絡み辛いだけや(笑)
それとバカッターを筆頭に
陽気であれば何でも出来る(キリッって国民性なのが日本人(笑)だからなw
つまらない芸人に法的措置を取るよと言われて
社会主義だ全体主義だと叫ぶ馬鹿日本人(笑)
日本って反日本主義や反日本人主義なだけだと思うw社会主義にすらなってない。
日本や日本人が嫌いなのは分かるが
標準(笑)と変わらないのが大半だろうな。
>>762
初期農耕民のY−DNAマーカーって結構分かってるの?
俺はここに出てるアイスマンぐらいしかしらない
Analysis of his mitochondrial DNA has shown that Otzi belongs to the K1 subclade, but cannot be categorized into any of the three modern branches of that subclade (K1a, K1b or K1c). The new subclade has provisionally been named K1o for Otzi.[32]
Multiplex assay study was able to confirm that the Iceman's mtDNA belongs to a new European mtDNA clade with a very limited distribution amongst modern data sets.[33]
初期農耕民のY−DNAマーカーって結構分かってるの?
俺はここに出てるアイスマンぐらいしかしらない
Analysis of his mitochondrial DNA has shown that Otzi belongs to the K1 subclade, but cannot be categorized into any of the three modern branches of that subclade (K1a, K1b or K1c). The new subclade has provisionally been named K1o for Otzi.[32]
Multiplex assay study was able to confirm that the Iceman's mtDNA belongs to a new European mtDNA clade with a very limited distribution amongst modern data sets.[33]
>>556>>557
BP7500頃 新石器文化(LBK)のH'グループが狩猟採集民のUグループに進入
BP6500頃 新石器文化(LBK)のH'グループが衰退し、現在の欧州人のハプロ構成に近くなる
ってことで、まずこれが前提
>>761そのうえで、この後期の変異はどこから持ち込まれたかって話だから
少なくとも3つ以上だよ
BP7500頃 新石器文化(LBK)のH'グループが狩猟採集民のUグループに進入
BP6500頃 新石器文化(LBK)のH'グループが衰退し、現在の欧州人のハプロ構成に近くなる
ってことで、まずこれが前提
>>761そのうえで、この後期の変異はどこから持ち込まれたかって話だから
少なくとも3つ以上だよ
>>764
すまんこれはmtDNAだった
>>753>>749 Yのハプロはこっち
The Y-DNA of Otzi belongs to a subclade of G defined by the SNPs M201, P287, P15, L223 and L91 (G-L91, ISOGG G2a2b, former "G2a4").
He was not typed for any of the subclades downstreaming from G-L91. G-L91 is now mostly found in South Corsica.
すまんこれはmtDNAだった
>>753>>749 Yのハプロはこっち
The Y-DNA of Otzi belongs to a subclade of G defined by the SNPs M201, P287, P15, L223 and L91 (G-L91, ISOGG G2a2b, former "G2a4").
He was not typed for any of the subclades downstreaming from G-L91. G-L91 is now mostly found in South Corsica.
>>757 凄い早期なんだよな
http://www.um.u-tokyo.ac.jp/publish_db/2007moundsAndGoodesses/07/007_01_01.html
■西アジアにおける有蹄類の家畜化
家畜化の起源地
ヒツジ、ヤギ、ウシ、ブタという有蹄類の家畜化は、肉食動物(イヌやネコ)の家畜化よりも遅れて始まったが、
食糧供給に大きな変化をもたらした点で、より重要である。有蹄類が最初に家畜化された時期と場所については、過去数十年間、新たな説が繰り返し提示されてきた。
前8千年紀のイランとする説からはじまり、前7千年紀のイスラエルとする説、前8千年紀のシリアとする説、
前9千年紀の南東アナトリア(トルコ)とする説まである(Harris 1996;Vigne et al. 2005など)。
現在では、ほとんどの考古学者が、最初の家畜化は前9千年紀半ば(PPNB前期)に、南東アナトリアのタウルス山脈南麓で始まったと考えている。
たとえば、トルコのネバル・チョリ遺跡のPPNB前期(前8,500年)層では、ヒツジ(Ovis orientalis)とヤギ(Capra aegagrus)の小型化が顕著になると同時に、
幼獣個体の比率が増加することが明らかとなっている(Peters et al. 2005)。
同時期のガゼルには形態変化は全く認められていないから、それは家畜化の結果であると考えられる(図1・2)。
また、ほぼ同時期の北シリア、ジャーデ遺跡でも、性的二形がやや小さくなったことがわかっている(Helmer et al. 2005)(図3)。
この現象も、家畜化開始に関する確実性の高い指標とされているものである。一方、ブタの家畜化はもっと漸移的に進行したようである(Erwinck et al. 2002)。
家畜は、その起源地である南東アナトリアから急速に広がっていった。
http://www.um.u-tokyo.ac.jp/publish_db/2007moundsAndGoodesses/07/007_01_01.html
■西アジアにおける有蹄類の家畜化
家畜化の起源地
ヒツジ、ヤギ、ウシ、ブタという有蹄類の家畜化は、肉食動物(イヌやネコ)の家畜化よりも遅れて始まったが、
食糧供給に大きな変化をもたらした点で、より重要である。有蹄類が最初に家畜化された時期と場所については、過去数十年間、新たな説が繰り返し提示されてきた。
前8千年紀のイランとする説からはじまり、前7千年紀のイスラエルとする説、前8千年紀のシリアとする説、
前9千年紀の南東アナトリア(トルコ)とする説まである(Harris 1996;Vigne et al. 2005など)。
現在では、ほとんどの考古学者が、最初の家畜化は前9千年紀半ば(PPNB前期)に、南東アナトリアのタウルス山脈南麓で始まったと考えている。
たとえば、トルコのネバル・チョリ遺跡のPPNB前期(前8,500年)層では、ヒツジ(Ovis orientalis)とヤギ(Capra aegagrus)の小型化が顕著になると同時に、
幼獣個体の比率が増加することが明らかとなっている(Peters et al. 2005)。
同時期のガゼルには形態変化は全く認められていないから、それは家畜化の結果であると考えられる(図1・2)。
また、ほぼ同時期の北シリア、ジャーデ遺跡でも、性的二形がやや小さくなったことがわかっている(Helmer et al. 2005)(図3)。
この現象も、家畜化開始に関する確実性の高い指標とされているものである。一方、ブタの家畜化はもっと漸移的に進行したようである(Erwinck et al. 2002)。
家畜は、その起源地である南東アナトリアから急速に広がっていった。
>>768
ヒツジとヤギがユーフラテス川中流域に出現するのは前8千年紀の初頭(PPNB中期)のことである。
テル・ハルーラやテル・アブ・フレイラ(Peters et al. 1999)などの遺跡でその存在が確認されている。
ユーフラテス中流域はヒツジやヤギの野生種が本来いなかった地域であるから、よそから持ち込まれたことは明らかである。
また、さらに遠方へ持ち込まれたこともわかっている。ダマスカス周辺地域(テル・アスワド、Helmer in press)はもとより、
海を越えてキプロス島(シルロカンボス、Vigne et al. 2000, 2003, in prep.)にまで至った。
PPNB後期末(前7,000年頃)に入ると、家畜は地中海沿岸からザグロス山脈まで、あるいはタウルス山脈からネゲヴ地方に至るあらゆる地域でみられるようになる。
ただし、こうした見方とは別に、家畜化の中心地が複数存在したという説もある。
例えば、ヤギの家畜化の中心地としてザグロス山脈西部(Zeder 2005)が、ウシに関してインダス川下流域(Meadow 1989)が想定されている。
しかし、そうした地域における家畜出現年代はやや新しく、それぞれ前8千年紀前半と前7千年紀という年代が得られている。
家畜化の開始という点では、やはり、南東アナトリアが初現と考えておいてよいだろう。
新石器化過程は、定住や植物栽培、動物家畜化などがからみあった現象であるが、動物の家畜化はその中ではかなり遅い段階で始まった(図4)。
部分的に定住が始まったナトゥーフ期からは3,000年、さらに、原初的な穀物栽培の出現よりも1,000年から1,500年ほど遅れて始まった。
つまり、動物の家畜化というのは、土器出現以前の新石器化現象のなかでは、最後の重大な技術経済的変化とみることができる(図4)。
ヒツジとヤギがユーフラテス川中流域に出現するのは前8千年紀の初頭(PPNB中期)のことである。
テル・ハルーラやテル・アブ・フレイラ(Peters et al. 1999)などの遺跡でその存在が確認されている。
ユーフラテス中流域はヒツジやヤギの野生種が本来いなかった地域であるから、よそから持ち込まれたことは明らかである。
また、さらに遠方へ持ち込まれたこともわかっている。ダマスカス周辺地域(テル・アスワド、Helmer in press)はもとより、
海を越えてキプロス島(シルロカンボス、Vigne et al. 2000, 2003, in prep.)にまで至った。
PPNB後期末(前7,000年頃)に入ると、家畜は地中海沿岸からザグロス山脈まで、あるいはタウルス山脈からネゲヴ地方に至るあらゆる地域でみられるようになる。
ただし、こうした見方とは別に、家畜化の中心地が複数存在したという説もある。
例えば、ヤギの家畜化の中心地としてザグロス山脈西部(Zeder 2005)が、ウシに関してインダス川下流域(Meadow 1989)が想定されている。
しかし、そうした地域における家畜出現年代はやや新しく、それぞれ前8千年紀前半と前7千年紀という年代が得られている。
家畜化の開始という点では、やはり、南東アナトリアが初現と考えておいてよいだろう。
新石器化過程は、定住や植物栽培、動物家畜化などがからみあった現象であるが、動物の家畜化はその中ではかなり遅い段階で始まった(図4)。
部分的に定住が始まったナトゥーフ期からは3,000年、さらに、原初的な穀物栽培の出現よりも1,000年から1,500年ほど遅れて始まった。
つまり、動物の家畜化というのは、土器出現以前の新石器化現象のなかでは、最後の重大な技術経済的変化とみることができる(図4)。
770 :世界@名無史さん:2014/01/22(水) 09:47:26.37 0
雌が本来無角で扱いやすそうな鹿より、両性ともに角があるウシ科を家畜化の対象に選んだのが不思議
771 :世界@名無史さん:2014/01/22(水) 10:21:50.46 0
鹿は乳が出ないだろう。しかも不味い
772 :世界@名無史さん:2014/01/22(水) 11:56:45.54 0
動画 You Tube ドイツ/移民政策の限界
http://www.youtube.com/watch?v=Rvb-7KfKPwA
動画 You Tube ノルウェー/多発する移民による強姦事件
http://www.youtube.com/watch?v=1LG-5QziOWo
http://www.youtube.com/watch?v=Rvb-7KfKPwA
動画 You Tube ノルウェー/多発する移民による強姦事件
http://www.youtube.com/watch?v=1LG-5QziOWo
>>762
Origins of the Etruscans: Was Herodotus right?
By Nicholas Wade
Published: Tuesday, April 3, 2007
http://www.nytimes.com/2007/04/03/health/03iht-snetrus.1.5127788.html?_r=1&
2007年 03月 16日
エトルリア人の起源ー牛のDNAにより解明?
http://jamarte.exblog.jp/4866896
エトルリア人の起源の新説についてのやや苦笑を誘う記事を読んだ。
エトルリア人がどこから来たのかを解明するのにあたり、
トスカーナ地方に特有の白いキアニーナ牛(かのビステッカ・アラ・フィオレンティーナの牛)のDNAが
いわゆるアナトリア半島や中近東の牛のDNAと一番近いというのが調査で明らかになったことから、
トスカーナ地方近辺に栄えたエトルリア文明の起源はここにあり、というのである。
500頭にわたるこの牛種を検査したところこのキアニーナ牛と一番近いDNAを持つのは地理的に近いヨーロッパの他の地域や北アフリカの牛ではなく、
アナトリア、中近東の牛であり、また牛に限らずトスカーナ地方の住民の5%のDNAが遺伝子学上中近東の人々のDNAに近いこともわかった。
特にそれが顕著なののはエトルリア人の街として有名なムルロ(Murlo)やカゼンティーノ(Casentino)の住民であるそうだ。
牛のDNAの類似点だけであるなら、当時商業的にかの地から持ち込まれたのではないか、という疑問も残ったのであるが、
人間のDNAもしかり、ということで、この時代にアナトリア地方から移住してきた民族がエトルリアの先祖であるという説が堅くなってきた。
それも陸地移住ではなく海から渡ってきたらしい。
陸地つたいであるなら必ず通るはずのトリエステやアルプス山脈沿いの街にこの遺伝子の種子の跡が全く発見されていないことがその理由である。
DNA Boosts Herodotus’ Account of Etruscans as Migrants to Italy
http://www.nytimes.com/2007/04/03/science/03etruscan.html?pagewanted=all
Origins of the Etruscans: Was Herodotus right?
By Nicholas Wade
Published: Tuesday, April 3, 2007
http://www.nytimes.com/2007/04/03/health/03iht-snetrus.1.5127788.html?_r=1&
2007年 03月 16日
エトルリア人の起源ー牛のDNAにより解明?
http://jamarte.exblog.jp/4866896
エトルリア人の起源の新説についてのやや苦笑を誘う記事を読んだ。
エトルリア人がどこから来たのかを解明するのにあたり、
トスカーナ地方に特有の白いキアニーナ牛(かのビステッカ・アラ・フィオレンティーナの牛)のDNAが
いわゆるアナトリア半島や中近東の牛のDNAと一番近いというのが調査で明らかになったことから、
トスカーナ地方近辺に栄えたエトルリア文明の起源はここにあり、というのである。
500頭にわたるこの牛種を検査したところこのキアニーナ牛と一番近いDNAを持つのは地理的に近いヨーロッパの他の地域や北アフリカの牛ではなく、
アナトリア、中近東の牛であり、また牛に限らずトスカーナ地方の住民の5%のDNAが遺伝子学上中近東の人々のDNAに近いこともわかった。
特にそれが顕著なののはエトルリア人の街として有名なムルロ(Murlo)やカゼンティーノ(Casentino)の住民であるそうだ。
牛のDNAの類似点だけであるなら、当時商業的にかの地から持ち込まれたのではないか、という疑問も残ったのであるが、
人間のDNAもしかり、ということで、この時代にアナトリア地方から移住してきた民族がエトルリアの先祖であるという説が堅くなってきた。
それも陸地移住ではなく海から渡ってきたらしい。
陸地つたいであるなら必ず通るはずのトリエステやアルプス山脈沿いの街にこの遺伝子の種子の跡が全く発見されていないことがその理由である。
DNA Boosts Herodotus’ Account of Etruscans as Migrants to Italy
http://www.nytimes.com/2007/04/03/science/03etruscan.html?pagewanted=all
>>775
Mitochondrial DNA Variation of Modern Tuscans Supports the Near Eastern Origin of Etruscans
http://www.sciencedirect.com/science/article/pii/S0002929707611069
Mitochondrial DNA Variation of Modern Tuscans Supports the Near Eastern Origin of Etruscans
http://www.sciencedirect.com/science/article/pii/S0002929707611069
777 :世界@名無史さん:2014/01/24(金) 02:59:27.12 0
ヨーロッパの牛は全て中東のオーロックスから家畜化されたはずなんだが。ヨーロッパにいたオーロックスは家畜の牛の遺伝子プールに殆ど寄与していないらしい
何波も家畜を伴った農耕民の移住があって、キアニーナ牛とその飼い主は後発組ってことか?
何波も家畜を伴った農耕民の移住があって、キアニーナ牛とその飼い主は後発組ってことか?
>>775これはこっちだった>>750
>>777
http://en.wikipedia.org/wiki/Aurochs
The aurochs, which ranged throughout much of Eurasia and Northern Africa during the late Pleistocene and early Holocene, is widely accepted as the wild ancestor of modern cattle.
Archaeological evidence shows that domestication occurred
independently in the Near East and the Indian subcontinent between 10,000?8,000 years ago,
giving rise to the two major domestic taxa observed today: humpless Bos taurus (taurine) and humped Bos indicus (zebu), respectively.
This is confirmed by genetic analyses of matrilineal mitochondrial DNA sequences, which reveal a marked differentiation between modern Bos taurus and Bos indicus haplotypes,
demonstrating their derivation from two geographically and genetically divergent wild populations.[11]
It is possible that there was a third domestication event from another form of the Aurochs in Africa.
この辺りは記憶と一致
>>777
http://en.wikipedia.org/wiki/Aurochs
The aurochs, which ranged throughout much of Eurasia and Northern Africa during the late Pleistocene and early Holocene, is widely accepted as the wild ancestor of modern cattle.
Archaeological evidence shows that domestication occurred
independently in the Near East and the Indian subcontinent between 10,000?8,000 years ago,
giving rise to the two major domestic taxa observed today: humpless Bos taurus (taurine) and humped Bos indicus (zebu), respectively.
This is confirmed by genetic analyses of matrilineal mitochondrial DNA sequences, which reveal a marked differentiation between modern Bos taurus and Bos indicus haplotypes,
demonstrating their derivation from two geographically and genetically divergent wild populations.[11]
It is possible that there was a third domestication event from another form of the Aurochs in Africa.
この辺りは記憶と一致
>>778続き
The Sanga cattle, a not-humped zebu like cattle type, is commonly believed to originate from crosses between humped-zebus with taurine cattle breeds.
However, there is archaeological evidence that these cattle were domesticated independently in Africa and that bloodlines of taurine and zebu cattle were introduced only within the last few hundreds years.[24]
Domestication of the aurochs began in the southern Caucasus and northern Mesopotamia from about the 6th millennium BC.
Genetic evidence suggests that aurochs were independently domesticated in India and possibly also in northern Africa.[25]
Domesticated cattle and aurochs are so different in size that they have been regarded as separate species;
however, large ancient cattle and aurochs "are difficult to classify because morphological traits have overlapping distributions in cattle and aurochs and diagnostic features are identified only in horn and some cranial element."[8][26]
A DNA study suggests that all domesticated taurine cattle originated from about 80 wild aurochs. Those animals lived in Iran 10,500 years ago.[27]
これも何処かでみかけた
でも、知ってる有名な遺跡ですぐに思いつくのは「チャタル・ヒュユク」
http://ja.wikipedia.org/wiki/%E3%83%81%E3%83%A3%E3%82%BF%E3%83%AB%E3%83%BB%E3%83%92%E3%83%A5%E3%83%A6%E3%82%AF
The Sanga cattle, a not-humped zebu like cattle type, is commonly believed to originate from crosses between humped-zebus with taurine cattle breeds.
However, there is archaeological evidence that these cattle were domesticated independently in Africa and that bloodlines of taurine and zebu cattle were introduced only within the last few hundreds years.[24]
Domestication of the aurochs began in the southern Caucasus and northern Mesopotamia from about the 6th millennium BC.
Genetic evidence suggests that aurochs were independently domesticated in India and possibly also in northern Africa.[25]
Domesticated cattle and aurochs are so different in size that they have been regarded as separate species;
however, large ancient cattle and aurochs "are difficult to classify because morphological traits have overlapping distributions in cattle and aurochs and diagnostic features are identified only in horn and some cranial element."[8][26]
A DNA study suggests that all domesticated taurine cattle originated from about 80 wild aurochs. Those animals lived in Iran 10,500 years ago.[27]
これも何処かでみかけた
でも、知ってる有名な遺跡ですぐに思いつくのは「チャタル・ヒュユク」
http://ja.wikipedia.org/wiki/%E3%83%81%E3%83%A3%E3%82%BF%E3%83%AB%E3%83%BB%E3%83%92%E3%83%A5%E3%83%A6%E3%82%AF
Modern Taurine Cattle Descended from Small Number of Near-Eastern Founders
http://mbe.oxfordjournals.org/content/29/9/2101.full
The earliest signs of wild aurochs domestication are seen at Dja'de in the Middle Euphrates Valley, dating to the Early Pre-Pottery Neolithic (EPPNB; 10,800?10,300 cal. BP, Helmer et al. 2005)
and at Cayonu in the High Tigris Valley, between the Early and Middle PPNB (around 10,200 cal. BP, Hongo et al. 2009).
After an initial breeding phase lasting some 1.5 millennia in an area between the Levant, central Anatolia and western Iran,
domestic cattle started to appear in western Anatolia and southeastern Europe by 8,800 cal. BP,
southern Italy by 8,500 cal. BP, and Central Europe by 8,000 cal. BP.
Archaeozoological (Helmer and Vigne 2007; Vigne 2008) residual lipid (Craig et al. 2005; Evershed et al. 2008)
and lactase persistence data (Itan et al. 2009) point to an increasing economic importance of cattle for meat and milk production.
ということみたいなので>>777そうなんじゃなかろうか
http://mbe.oxfordjournals.org/content/29/9/2101.full
The earliest signs of wild aurochs domestication are seen at Dja'de in the Middle Euphrates Valley, dating to the Early Pre-Pottery Neolithic (EPPNB; 10,800?10,300 cal. BP, Helmer et al. 2005)
and at Cayonu in the High Tigris Valley, between the Early and Middle PPNB (around 10,200 cal. BP, Hongo et al. 2009).
After an initial breeding phase lasting some 1.5 millennia in an area between the Levant, central Anatolia and western Iran,
domestic cattle started to appear in western Anatolia and southeastern Europe by 8,800 cal. BP,
southern Italy by 8,500 cal. BP, and Central Europe by 8,000 cal. BP.
Archaeozoological (Helmer and Vigne 2007; Vigne 2008) residual lipid (Craig et al. 2005; Evershed et al. 2008)
and lactase persistence data (Itan et al. 2009) point to an increasing economic importance of cattle for meat and milk production.
ということみたいなので>>777そうなんじゃなかろうか
>>780
Dja'de Early Pre-Pottery Neolithic (EPPNB; 10,800?10,300 cal. BP, Helmer et al. 2005)
Cayonu Early and Middle PPNB (around 10,200 cal. BP, Hongo et al. 2009).
western Anatolia 8,800 cal. BP,
southeastern Europe 8,800 cal. BP,
southern Italy by 8,500 cal. BP
Central Europe by 8,000 cal. BP
>>757
ポーランド ボグスキは新石器時代 紀元前5,500 7,500BP
たしかデンマークもかなり早かった
Dja'de Early Pre-Pottery Neolithic (EPPNB; 10,800?10,300 cal. BP, Helmer et al. 2005)
Cayonu Early and Middle PPNB (around 10,200 cal. BP, Hongo et al. 2009).
western Anatolia 8,800 cal. BP,
southeastern Europe 8,800 cal. BP,
southern Italy by 8,500 cal. BP
Central Europe by 8,000 cal. BP
>>757
ポーランド ボグスキは新石器時代 紀元前5,500 7,500BP
たしかデンマークもかなり早かった
782 :世界@名無史さん:2014/01/24(金) 22:16:53.80 0
寒いところでチーズ?とかいう人がいるけど、当時のポーランドは今よりかなり温暖だった
日本だと縄文海進の時期だし
日本だと縄文海進の時期だし
783 :世界@名無史さん:2014/01/24(金) 22:18:59.51 0
つまり完新世気候最温暖期
暖かいポーランドでいまのギリシャのフェタチーズのような白チーズを山羊乳で作って食べていた
暖かいポーランドでいまのギリシャのフェタチーズのような白チーズを山羊乳で作って食べていた
784 :世界@名無史さん:2014/01/24(金) 22:21:03.43 0
イタリア人はR1bの比率が意外と低い。
印欧語族の先住民の子孫が今でもかなりの割合になると言う意味だね。
印欧語族の先住民の子孫が今でもかなりの割合になると言う意味だね。
785 :世界@名無史さん:2014/01/24(金) 22:28:49.28 0
ヨーロッパ北部(アルプス〜カルパチア以北)の先住民つまり狩猟採集民はウラル系だろう。Iなど。
そこへやってきて互いに嫌いあっていた農耕民はプレ印欧語族の言語の話し手でこれはR1b。
その後も何度か小さな移民の波があり、その時代に東方でR1bとR1aの分化が進んだ。
R1aはステップ地帯に進出し移牧ないし遊牧の技術を身に着けた。
彼らは牧畜民として父系社会も形成する。また階層分化も大きくなり墳墓(クルガン)の文化を作る。
プロト印欧語の人々だ。
そして彼らが最初に大規模に欧州に侵入した形跡である考古文化が「球状アンフォラ文化」。
そこへやってきて互いに嫌いあっていた農耕民はプレ印欧語族の言語の話し手でこれはR1b。
その後も何度か小さな移民の波があり、その時代に東方でR1bとR1aの分化が進んだ。
R1aはステップ地帯に進出し移牧ないし遊牧の技術を身に着けた。
彼らは牧畜民として父系社会も形成する。また階層分化も大きくなり墳墓(クルガン)の文化を作る。
プロト印欧語の人々だ。
そして彼らが最初に大規模に欧州に侵入した形跡である考古文化が「球状アンフォラ文化」。
786 :世界@名無史さん:2014/01/24(金) 22:31:49.99 0
イタリアなど欧州南部は地中海を行き来できるため地中海世界に面した人々が混在していく。
そのためアルプス〜カルパチア以南の諸国(南部諸国)の地域は北部よりもハプロタイプのバラエティが多い。
アルプス〜カルパチア以北の諸国(北部諸国)の地域ではせいぜいウラル系とプレ印欧語系とプロト印欧語系と印欧語のインド・イラン系の影響しか受けていない。
そのためアルプス〜カルパチア以南の諸国(南部諸国)の地域は北部よりもハプロタイプのバラエティが多い。
アルプス〜カルパチア以北の諸国(北部諸国)の地域ではせいぜいウラル系とプレ印欧語系とプロト印欧語系と印欧語のインド・イラン系の影響しか受けていない。
もともと欧州全域にプロト・ウラル語の人々が狩猟採集民として広く薄く住んでいたんじゃないのか?
とりあえず俺の見立てとして落ちるのはレムノス島
>>750
沿革[編集]
ヘロドトスによれば、エトルリア人は小アジアのリディアからこの地にやってきたと言う。
一方、ハリカルナッソスのディオニュシオスは、エトルリア人はイタリア古来の民族だと述べている。
現在の調査では正式には、エトルリア人が小アジアの出自である事を直接結び付ける証拠はない。
しかしながら、ある調査ではエーゲ海のレムノス島では紀元前6世紀までインド・ヨーロッパ語ではない民族が居住していた跡が見られ、その民族の言語がエトルリア人と似ている事が指摘されている。
また、エトルリア人は海を往来する民族でもあり、古代地中海世界の至るところからその存在が記述されている。一説には古代エジプト第20王朝に記述のある「海の民」はエトルリア人ではなかったかとも言われている。
>>616
>>750
沿革[編集]
ヘロドトスによれば、エトルリア人は小アジアのリディアからこの地にやってきたと言う。
一方、ハリカルナッソスのディオニュシオスは、エトルリア人はイタリア古来の民族だと述べている。
現在の調査では正式には、エトルリア人が小アジアの出自である事を直接結び付ける証拠はない。
しかしながら、ある調査ではエーゲ海のレムノス島では紀元前6世紀までインド・ヨーロッパ語ではない民族が居住していた跡が見られ、その民族の言語がエトルリア人と似ている事が指摘されている。
また、エトルリア人は海を往来する民族でもあり、古代地中海世界の至るところからその存在が記述されている。一説には古代エジプト第20王朝に記述のある「海の民」はエトルリア人ではなかったかとも言われている。
>>616
エトルリア語、ハンガリー語、トルコ語、似ている
バスク語とウラル語派諸語も似てるっちゃ似てるんだよな
798 :世界@名無史さん:2014/01/25(土) 07:24:02.20 0
遺伝子研究のおかげで欧州古代史がより鮮明になってきている。
少なくともクロマニヨン人の子孫という主張は影をひそめた。
印欧語族と遺伝子と酪農と人種のほぼ四身一体が公認されるまでもう少しだな。
少なくともクロマニヨン人の子孫という主張は影をひそめた。
印欧語族と遺伝子と酪農と人種のほぼ四身一体が公認されるまでもう少しだな。
ウラル系っていうか、先ウラル系(プロトウラリック)からの系譜だろうよIは
その現存子孫がウラル語派諸民族やバルカン西部諸民族など
バスクは先印欧語だろうよR1b
プレプロトインドヨーロピアン
この農耕民が、先ウラル系狩猟民が広く薄く住んでいた欧州全体に広まった
互いに嫌いあっていたようだが(人種差別)Iの容貌はコミ人のように白く薄く丸い顔金髪碧眼だろう
R1bになると鋭角的で精悍な、ただし少し浅黒い金髪碧眼で食糧事情が良い事で背が高い
R1bからするとIはドワーフに見えただろう
印欧語が成立したのはこの最初の農耕民が定住した後はるか東方の地で
この時代東方でR1bのほかR1aが成立
R1aはヤームナヤ文化やドニエプル・ドネツ文化の地で発生しただろう
ヤームナヤ文化はスキタイなどイラン系の文化に似ているが当時はまだケントゥム語だったろう
その現存子孫がウラル語派諸民族やバルカン西部諸民族など
バスクは先印欧語だろうよR1b
プレプロトインドヨーロピアン
この農耕民が、先ウラル系狩猟民が広く薄く住んでいた欧州全体に広まった
互いに嫌いあっていたようだが(人種差別)Iの容貌はコミ人のように白く薄く丸い顔金髪碧眼だろう
R1bになると鋭角的で精悍な、ただし少し浅黒い金髪碧眼で食糧事情が良い事で背が高い
R1bからするとIはドワーフに見えただろう
印欧語が成立したのはこの最初の農耕民が定住した後はるか東方の地で
この時代東方でR1bのほかR1aが成立
R1aはヤームナヤ文化やドニエプル・ドネツ文化の地で発生しただろう
ヤームナヤ文化はスキタイなどイラン系の文化に似ているが当時はまだケントゥム語だったろう
要はクルガン仮説に沿った説明だがね
そういえばこれの中身を確認するの忘れてた
【生物】家畜馬の祖先16万年前に急増、北米からアジア東部へ移動後 英など国際研究
http://anago.2ch.net/test/read.cgi/scienceplus/1336663217/1
1 名前:一般人φ ★[] 投稿日:2012/05/11(金) 00:20:17.62 ID:???
家畜の馬の祖先種は約16万年前にアジア東部で急増し、約5500年前までにカザフスタンやロシア南西部、
ウクライナ付近で家畜化された可能性が高いことが分かった。英ケンブリッジ大などの国際研究チームが
ユーラシア大陸各地の約300頭の馬から採取された細胞核DNAを解析した成果で、9日までに米科学アカデミー
紀要電子版に発表した。 これまでの研究によると、祖先種はもともと北米大陸で出現し、昔は陸続きだった
ベーリング海峡を経由して20万年前までにアジアに移動したとされていた。カザフスタン北部の遺跡で見つかった
約5500年前の馬の歯にはくつわをかませた跡があったり、陶器に馬乳のかすが付着したりしており、
今回の解析結果と一致するという。
▽PNAS
「Reconstructing the origin and spread of horse domestication in the Eurasian steppe」
http://www.pnas.org/content/early/2012/05/02/1111122109.abstract
【生物】家畜馬の祖先16万年前に急増、北米からアジア東部へ移動後 英など国際研究
http://anago.2ch.net/test/read.cgi/scienceplus/1336663217/1
1 名前:一般人φ ★[] 投稿日:2012/05/11(金) 00:20:17.62 ID:???
家畜の馬の祖先種は約16万年前にアジア東部で急増し、約5500年前までにカザフスタンやロシア南西部、
ウクライナ付近で家畜化された可能性が高いことが分かった。英ケンブリッジ大などの国際研究チームが
ユーラシア大陸各地の約300頭の馬から採取された細胞核DNAを解析した成果で、9日までに米科学アカデミー
紀要電子版に発表した。 これまでの研究によると、祖先種はもともと北米大陸で出現し、昔は陸続きだった
ベーリング海峡を経由して20万年前までにアジアに移動したとされていた。カザフスタン北部の遺跡で見つかった
約5500年前の馬の歯にはくつわをかませた跡があったり、陶器に馬乳のかすが付着したりしており、
今回の解析結果と一致するという。
▽PNAS
「Reconstructing the origin and spread of horse domestication in the Eurasian steppe」
http://www.pnas.org/content/early/2012/05/02/1111122109.abstract
>カザフスタンやロシア南西部、ウクライナ付近
まぎれもなくスレドニ・ストグ文化とフヴァリンスク文化だ…
まぎれもなくスレドニ・ストグ文化とフヴァリンスク文化だ…
>>781
上の二つのPPNBの真ん中がギョベクリ・テペ
http://ja.wikipedia.org/wiki/%E3%82%AE%E3%83%A7%E3%83%99%E3%82%AF%E3%83%AA%E3%83%BB%E3%83%86%E3%83%9A
そんでLBKとは年代が合わない気がするんだよな
Linear Pottery culture
http://en.wikipedia.org/wiki/Linear_Pottery_culture
The Linear Pottery culture is a major archaeological horizon of the European Neolithic, flourishing circa 5500?4500 BC.
It is abbreviated as LBK (from German: Linearbandkeramik),
and is also known as the Linear Band Ware, Linear Ware, Linear Ceramics or Incised Ware culture,
and falls within the Danubian I culture of V. Gordon Childe.
上の二つのPPNBの真ん中がギョベクリ・テペ
http://ja.wikipedia.org/wiki/%E3%82%AE%E3%83%A7%E3%83%99%E3%82%AF%E3%83%AA%E3%83%BB%E3%83%86%E3%83%9A
そんでLBKとは年代が合わない気がするんだよな
Linear Pottery culture
http://en.wikipedia.org/wiki/Linear_Pottery_culture
The Linear Pottery culture is a major archaeological horizon of the European Neolithic, flourishing circa 5500?4500 BC.
It is abbreviated as LBK (from German: Linearbandkeramik),
and is also known as the Linear Band Ware, Linear Ware, Linear Ceramics or Incised Ware culture,
and falls within the Danubian I culture of V. Gordon Childe.
>>781
Cardium Pottery
http://en.wikipedia.org/wiki/Cardium_Pottery
Preboreal (10.3 ka ? 9 ka),
Boreal (9 ka ? 7.5 ka),
Atlantic (7.5 ka ? 5 ka),
Subboreal (5 ka ? 2.5 ka)
Subatlantic (2.5 ka ? present)
http://upload.wikimedia.org/wikipedia/commons/8/81/Cardial_map.png
The Mediterranean Neolithic[edit]
This pottery style gives its name to the main culture of the Mediterranean Neolithic: Cardium Pottery Culture or Cardial Culture, or Impressed Ware Culture,
which eventually extended from the Adriatic sea to the Atlantic coasts of Portugal and south to Morocco.[4]
The earliest Impressed Ware sites, dating to 6400-6200 BC, are in Epirus and Corfu.
Settlements then appear in Albania and Dalmatia on the eastern Adriatic coast dating to between 6100 and 5900 BC.[5]
The earliest date in Italy comes from Coppa Nevigata on the Adriatic coast of southern Italy, perhaps as early as 6000 cal B.C.
Also during Su Carroppu civilization in Sardinia, already in its early stages (low strata into Su Coloru cave, c. 6000 BC) early examples of cardial pottery appear.[6]
Northward and westward all secure radiocarbon dates are identical to those for Iberia c. 5500 cal B.C.,
which indicates a rapid spread of Cardial and related cultures: 2,000 km from the gulf of Genoa to the estuary of the Mondego in probably no more than 100?200 years. This suggests a seafaring expansion by planting colonies along the coast. [7]
Cardium Pottery
http://en.wikipedia.org/wiki/Cardium_Pottery
Preboreal (10.3 ka ? 9 ka),
Boreal (9 ka ? 7.5 ka),
Atlantic (7.5 ka ? 5 ka),
Subboreal (5 ka ? 2.5 ka)
Subatlantic (2.5 ka ? present)
http://upload.wikimedia.org/wikipedia/commons/8/81/Cardial_map.png
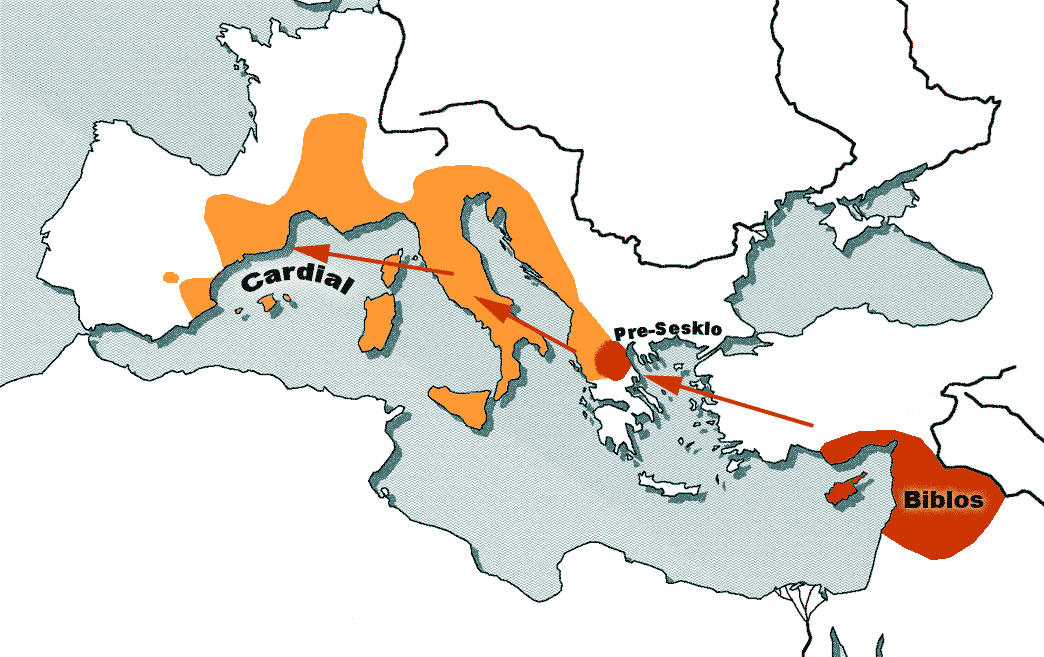{kind=link}
The Mediterranean Neolithic[edit]
This pottery style gives its name to the main culture of the Mediterranean Neolithic: Cardium Pottery Culture or Cardial Culture, or Impressed Ware Culture,
which eventually extended from the Adriatic sea to the Atlantic coasts of Portugal and south to Morocco.[4]
The earliest Impressed Ware sites, dating to 6400-6200 BC, are in Epirus and Corfu.
Settlements then appear in Albania and Dalmatia on the eastern Adriatic coast dating to between 6100 and 5900 BC.[5]
The earliest date in Italy comes from Coppa Nevigata on the Adriatic coast of southern Italy, perhaps as early as 6000 cal B.C.
Also during Su Carroppu civilization in Sardinia, already in its early stages (low strata into Su Coloru cave, c. 6000 BC) early examples of cardial pottery appear.[6]
Northward and westward all secure radiocarbon dates are identical to those for Iberia c. 5500 cal B.C.,
which indicates a rapid spread of Cardial and related cultures: 2,000 km from the gulf of Genoa to the estuary of the Mondego in probably no more than 100?200 years. This suggests a seafaring expansion by planting colonies along the coast. [7]
>>769
3) 無陶器・新石器文化/シロウロカンボス居住地遺跡
http://hp1.cyberstation.ne.jp/legend-ej/p-civil1997cyp-khir.html
パレクリシア村の北西1.5km、標高180mのポイントで1991年〜2004年までミッションを行ったフランス考古学チームは、
紀元前8500年〜前7000年頃に属する無陶器・新石器文化のシロウロカンボス居住地遺跡 Shillourokambos を発掘した。遺跡は海岸から約4kmの距離である。
この継続的な発掘ミッションでは、細い編み枝と粘土を使った泥漆喰で表層仕上げした「輪切りダイコン」のような形容の円筒形の家屋群、集落の中心には井戸も確認された。
また、出土品では黒曜石のフリント石器を初め、顔をモチーフにした石製像、装飾の小石、そして色々な動物の骨、埋葬人骨などが確認された。家屋構造や出土品に関しては、キロキティアの新石器文化と連動しているとされる。
2004年、フランス考古学チームの発表した発掘レポートでは、石器以外に大量のヒツジ、野生のロバやウシ、さらにイヌの仲間やイノシシ(ブタ)の骨まで出土したことから、このうち幾らかの動物は食料にするために家畜化されていたとされる。
これらの出土品はシロウロカンボスの人々が、野生動物の家畜化や種を蒔く定住した初期農業を営み、この地方の新石器文化のセンターであったキロキティア居住地と同様に、豊かで発達した生活を営んでいたことを暗示させている。
装飾石や磨製石斧、黒曜石のフリント石器類などが一緒に副葬されていたことから、かなり「地位」の高い人物であったことを推量させる30歳前後の屈葬人骨が見つかった。
興味深いことに、その屈葬人骨から40cm離れた地中から「埋葬された」と判断できるネコの完全骨格も見つかっている。
ネコの骨格発掘に関してフランス・チームの担当者は、紀元前7500年頃、今から約9,500年前の新石器文化の時代、シロウロカンボス居住地ではすでにネコを家庭内で飼う、いわゆる「飼いネコ/ペット」が行われていた、と判断している。
なお、シロウロカンボス居住地の「ある家族」が可愛がって飼っていた後、死亡したことから手厚く「埋葬した」と推測されるこのネコの骨格は、アフリカン・キャットの仲間で生後8か月〜9か月と推測されている。
3) 無陶器・新石器文化/シロウロカンボス居住地遺跡
http://hp1.cyberstation.ne.jp/legend-ej/p-civil1997cyp-khir.html
パレクリシア村の北西1.5km、標高180mのポイントで1991年〜2004年までミッションを行ったフランス考古学チームは、
紀元前8500年〜前7000年頃に属する無陶器・新石器文化のシロウロカンボス居住地遺跡 Shillourokambos を発掘した。遺跡は海岸から約4kmの距離である。
この継続的な発掘ミッションでは、細い編み枝と粘土を使った泥漆喰で表層仕上げした「輪切りダイコン」のような形容の円筒形の家屋群、集落の中心には井戸も確認された。
また、出土品では黒曜石のフリント石器を初め、顔をモチーフにした石製像、装飾の小石、そして色々な動物の骨、埋葬人骨などが確認された。家屋構造や出土品に関しては、キロキティアの新石器文化と連動しているとされる。
2004年、フランス考古学チームの発表した発掘レポートでは、石器以外に大量のヒツジ、野生のロバやウシ、さらにイヌの仲間やイノシシ(ブタ)の骨まで出土したことから、このうち幾らかの動物は食料にするために家畜化されていたとされる。
これらの出土品はシロウロカンボスの人々が、野生動物の家畜化や種を蒔く定住した初期農業を営み、この地方の新石器文化のセンターであったキロキティア居住地と同様に、豊かで発達した生活を営んでいたことを暗示させている。
装飾石や磨製石斧、黒曜石のフリント石器類などが一緒に副葬されていたことから、かなり「地位」の高い人物であったことを推量させる30歳前後の屈葬人骨が見つかった。
興味深いことに、その屈葬人骨から40cm離れた地中から「埋葬された」と判断できるネコの完全骨格も見つかっている。
ネコの骨格発掘に関してフランス・チームの担当者は、紀元前7500年頃、今から約9,500年前の新石器文化の時代、シロウロカンボス居住地ではすでにネコを家庭内で飼う、いわゆる「飼いネコ/ペット」が行われていた、と判断している。
なお、シロウロカンボス居住地の「ある家族」が可愛がって飼っていた後、死亡したことから手厚く「埋葬した」と推測されるこのネコの骨格は、アフリカン・キャットの仲間で生後8か月〜9か月と推測されている。
>>801
これは見れる
A High Density SNP Array for the Domestic Horse and Extant Perissodactyla: Utility for Association Mapping, Genetic Diversity, and Phylogeny Studies
http://www.plosgenetics.org/article/info%3Adoi%2F10.1371%2Fjournal.pgen.1002451
これは見れない
Evidence of Positive Selection for a Glycogen Synthase (GYS1) Mutation in Domestic Horse Populations
http://jhered.oxfordjournals.org/content/early/2013/11/07/jhered.est075.abstract
これは見れる
A High Density SNP Array for the Domestic Horse and Extant Perissodactyla: Utility for Association Mapping, Genetic Diversity, and Phylogeny Studies
http://www.plosgenetics.org/article/info%3Adoi%2F10.1371%2Fjournal.pgen.1002451
これは見れない
Evidence of Positive Selection for a Glycogen Synthase (GYS1) Mutation in Domestic Horse Populations
http://jhered.oxfordjournals.org/content/early/2013/11/07/jhered.est075.abstract
808 :世界@名無史さん:2014/01/26(日) 00:48:51.93 0
imgur.com/Hr4vyL7
ロリコンネットストーカーで有名なおっさん HN IGZYゆたんぽ
1963年1月21日生 相模高校1981年卒五十嵐良裕
この画像を見る限り 土人にしか見えない
ロリコンネットストーカーで有名なおっさん HN IGZYゆたんぽ
1963年1月21日生 相模高校1981年卒五十嵐良裕
この画像を見る限り 土人にしか見えない
南欧における初期農耕民とその他北部全体における初期農耕民は出自の異なる集団では?
811 :世界@名無史さん:2014/01/26(日) 12:43:15.74 0
2000年ぐらい前までは、北欧でも南欧系遺伝子の割合が高かったそうだ。
Iはともかく、R系遺伝子が欧州を覆うようになったのは意外と最近のことかもね。
Iはともかく、R系遺伝子が欧州を覆うようになったのは意外と最近のことかもね。
北部欧州の初期農耕民というのは考古文化区分で言うと何?
南欧出身だとしたらどこから?
南欧出身だとしたらどこから?
813 :世界@名無史さん:2014/01/26(日) 13:10:11.23 0
スタルチェヴォ文化(バルカン)→線帯文土器文化(LBK、北部欧州)
当時は東部でククテニ・トリポリエ文化も並立
当時は東部でククテニ・トリポリエ文化も並立
815 :世界@名無史さん:2014/01/26(日) 13:16:07.25 0
アトキンソン等はこれを初期印欧語文化だと決めつけたが、実際は違うな
このオッサンは考古遺物を無視しすぎ
このオッサンは考古遺物を無視しすぎ
817 :世界@名無史さん:2014/01/26(日) 13:26:21.65 0
>>816
ウクライナ方面から来てカルパチア山脈を境に二手に分かれた
北部方向は球状アンフォラ文化(北部欧州方面)
南部方向はヴィンチャ文化(南部欧州方面)
ヴィンチャ文化は俗に原ギリシャ人と呼ばれ、まずドーリア人がギリシャに入った
ウクライナ方面から来てカルパチア山脈を境に二手に分かれた
北部方向は球状アンフォラ文化(北部欧州方面)
南部方向はヴィンチャ文化(南部欧州方面)
ヴィンチャ文化は俗に原ギリシャ人と呼ばれ、まずドーリア人がギリシャに入った
818 :世界@名無史さん:2014/01/26(日) 13:29:11.10 0
表現が正しくなかった
南部方向はヴィンチャ文化の地域に侵入した、とするのが正しい
南部方向はヴィンチャ文化の地域に侵入した、とするのが正しい
すみません
ヴィンチャ文化じゃなくてスタルチェヴォ文化でしたね
ヴィンチャ文化じゃなくてスタルチェヴォ文化でしたね
>>815
スタルチェヴォ文化は謎だー
アナトリア仮説が示すように印欧語なのか
それともクルガン仮説が示すように非印欧語なのか
アナトリア仮説はかなり無理があるようにみえる
考古遺物の無視とはそういうことですね
個人的にはクルガン仮説を支持
スタルチェヴォ文化は謎だー
アナトリア仮説が示すように印欧語なのか
それともクルガン仮説が示すように非印欧語なのか
アナトリア仮説はかなり無理があるようにみえる
考古遺物の無視とはそういうことですね
個人的にはクルガン仮説を支持
>>660
ちなみに北欧とは言わないと思うが
ドイツの個体はLBKで、これも南欧系列
“Stuttgart”, a ~7,500 year old individual
found in Stuttgart in southern Germany who was buried in the context of artifacts from the first
widespread Neolithic farming culture of central and northern Europe, the Linearbandkeramik (LBK).
そんでもって俺は歴史区分も詳しくない
ちなみに北欧とは言わないと思うが
ドイツの個体はLBKで、これも南欧系列
“Stuttgart”, a ~7,500 year old individual
found in Stuttgart in southern Germany who was buried in the context of artifacts from the first
widespread Neolithic farming culture of central and northern Europe, the Linearbandkeramik (LBK).
そんでもって俺は歴史区分も詳しくない
824 :世界@名無史さん:2014/01/26(日) 13:51:40.30 0
余談だが>>817で言及した原ギリシャ人はヴチェドル文化(Vucedol)の後継のヴァテイン文化(vatin)
ここからドーリア人がまずギリシャに入った
ここからドーリア人がまずギリシャに入った
825 :世界@名無史さん:2014/01/26(日) 13:54:15.14 0
ゲルマン=スラヴ=バルト連続体
そろそろ次の引越し先を探していたりするけど
朝鮮系ばっかりでうんざりする
朝鮮系ばっかりでうんざりする
828 :世界@名無史さん:2014/01/26(日) 17:02:29.57 0
まあ、クルガン仮説が当たらずといえども遠からずだろうね。
>>804
6400-6200 BC, are in Epirus and Corfu
6100-5900 BC, Albania and Dalmatia on the eastern Adriatic coast
6000 cal B.C(as early as ) Italy comes from Coppa Nevigata on the Adriatic coast of southern Italy
c. 6000 BC Su Carroppu civilization in Sardinia,
c. 5500 cal B.C Iberia
>>781
500年ぐらいづつこっちのが早いけど、これも海上の道だろうな
6400-6200 BC, are in Epirus and Corfu
6100-5900 BC, Albania and Dalmatia on the eastern Adriatic coast
6000 cal B.C(as early as ) Italy comes from Coppa Nevigata on the Adriatic coast of southern Italy
c. 6000 BC Su Carroppu civilization in Sardinia,
c. 5500 cal B.C Iberia
>>781
500年ぐらいづつこっちのが早いけど、これも海上の道だろうな
>>762
おもしろかったです
ちなみに、これはどうしてだと思います?
http://en.wikipedia.org/wiki/Haplogroup_T-M184
Estonians Estonian (Uralic) Estonia 11/207 5.3%
ノイズではありえない数値に見えますが、、、
おもしろかったです
ちなみに、これはどうしてだと思います?
http://en.wikipedia.org/wiki/Haplogroup_T-M184
Estonians Estonian (Uralic) Estonia 11/207 5.3%
ノイズではありえない数値に見えますが、、、
>>830
Migration Waves to the Baltic Sea Region
http://onlinelibrary.wiley.com/doi/10.1111/j.1469-1809.2007.00429.x/full
ちと古い、、、
Migration Waves to the Baltic Sea Region
http://onlinelibrary.wiley.com/doi/10.1111/j.1469-1809.2007.00429.x/full
ちと古い、、、
833 :世界@名無史さん:2014/01/27(月) 19:21:48.25 0
>>829
この連中がE・J2かな。
この連中がE・J2かな。
834 :世界@名無史さん:2014/01/27(月) 21:57:14.90 0
ヒトラー改宗ユダヤ人子孫説が誤りかも知れないってことか。その時代にEがヨーロッパに入ってたとすると
7000年前頃のヨーロッパ人のゲノム
この研究では、スペインのレオンにあるラブラナアリンテーロ(La Brana-Arintero)遺跡で発見された7000年前頃の中石器時代の男性の歯からDNAが抽出・解析され、
アルプスで発見された5300年前頃の通称「アイスマン」など新石器時代のヨーロッパ人、
スウェーデン・フィンランド・シベリアの狩猟採集民の墓から入手した部分的な遺伝子標本、
現代ヨーロッパ人35名のDNAなどと比較されました。
その結果、上部旧石器時代から中石器時代の西部・中央部ユーラシアにわたる共通の古代ゲノムが識別されました。
また、現代ヨーロッパ人の遺伝子において病原体抵抗と関連する多くの派生的・適応的異型が、
すでに中石器時代のヨーロッパの狩猟採集民に存在していたことも明らかになりました。
このことから、ヨーロッパにおける旧石器時代からの遺伝的連続性が推測されます。もちろん、新石器時代以降も、ヨーロッパへの人々の移住はありましたが。
また、中石器時代ラブラナアリンテーロ人男性のいくつかの肌の色素形成遺伝子には、古代から継承されていた肌の色を濃くするような対立遺伝子が確認されました。
このことから、中石器時代のヨーロッパにおいて、薄い色の肌はまだ広まっていなかったのではないか、とこの研究は示唆しています。
ヨーロッパで薄い色の肌が広まったのは、農耕への依存度が強くなっていったことと関係があるのかもしれません。
高緯度地帯において穀物への依存を強めていく(生肉の消費量が減る)と、肌の色が薄い方がビタミンD不足に陥りにくくなります。
もっとも、それは関係なく、性選択か単なる偶然だった可能性も考えられますが。
参考文献:
Olalde I. et al.(2014): Derived immune and ancestral pigmentation alleles in a 7,000-year-old Mesolithic European. Nature.
http://dx.doi.org/10.1038/nature12960
この研究では、スペインのレオンにあるラブラナアリンテーロ(La Brana-Arintero)遺跡で発見された7000年前頃の中石器時代の男性の歯からDNAが抽出・解析され、
アルプスで発見された5300年前頃の通称「アイスマン」など新石器時代のヨーロッパ人、
スウェーデン・フィンランド・シベリアの狩猟採集民の墓から入手した部分的な遺伝子標本、
現代ヨーロッパ人35名のDNAなどと比較されました。
その結果、上部旧石器時代から中石器時代の西部・中央部ユーラシアにわたる共通の古代ゲノムが識別されました。
また、現代ヨーロッパ人の遺伝子において病原体抵抗と関連する多くの派生的・適応的異型が、
すでに中石器時代のヨーロッパの狩猟採集民に存在していたことも明らかになりました。
このことから、ヨーロッパにおける旧石器時代からの遺伝的連続性が推測されます。もちろん、新石器時代以降も、ヨーロッパへの人々の移住はありましたが。
また、中石器時代ラブラナアリンテーロ人男性のいくつかの肌の色素形成遺伝子には、古代から継承されていた肌の色を濃くするような対立遺伝子が確認されました。
このことから、中石器時代のヨーロッパにおいて、薄い色の肌はまだ広まっていなかったのではないか、とこの研究は示唆しています。
ヨーロッパで薄い色の肌が広まったのは、農耕への依存度が強くなっていったことと関係があるのかもしれません。
高緯度地帯において穀物への依存を強めていく(生肉の消費量が減る)と、肌の色が薄い方がビタミンD不足に陥りにくくなります。
もっとも、それは関係なく、性選択か単なる偶然だった可能性も考えられますが。
参考文献:
Olalde I. et al.(2014): Derived immune and ancestral pigmentation alleles in a 7,000-year-old Mesolithic European. Nature.
http://dx.doi.org/10.1038/nature12960
ほいきた
>>836
http://www.nature.com/nature/journal/vaop/ncurrent/fig_tab/nature12960_F1.html
そんで
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2735096/figure/F2/
>>836
http://www.nature.com/nature/journal/vaop/ncurrent/fig_tab/nature12960_F1.html
そんで
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2735096/figure/F2/
>>837
とりあえずフィンランド人だけ別もの
解析は、5体の古人骨と、La Brana1を分離するもの
結果は、初期農耕民と狩猟採取民の対比を示す
図をアバウトに見る限り>>819
>>697の 'Gok4' はテッサリアと関係があり
>>764>>767 Iceman はイタリアと関係が深い
とりあえずフィンランド人だけ別もの
解析は、5体の古人骨と、La Brana1を分離するもの
結果は、初期農耕民と狩猟採取民の対比を示す
図をアバウトに見る限り>>819
>>697の 'Gok4' はテッサリアと関係があり
>>764>>767 Iceman はイタリアと関係が深い
>>837
今のところ、あんまり見るべきものがない
アイルランドが最も左よりなのは
農耕の進入が遅れたからなのかどうかってぐらいっすか
スコットランドとデンマーク
それとスカンジナビアの少数民族に
焦点を当てたらいいかとおもうんだけど
こっちの>>656の本番の方に期待ですかね
あ、それとおんなじ図に>>693のLoschbourは載っけるべき
今のところ、あんまり見るべきものがない
アイルランドが最も左よりなのは
農耕の進入が遅れたからなのかどうかってぐらいっすか
スコットランドとデンマーク
それとスカンジナビアの少数民族に
焦点を当てたらいいかとおもうんだけど
こっちの>>656の本番の方に期待ですかね
あ、それとおんなじ図に>>693のLoschbourは載っけるべき
とりあえずこれ>>837の結果は
>>714を支持しているように見える
これだと何にも面白くなさそうなんで
>>830のジェファーソンのTは海路はるばる北方へ入植したとしたかったが
>>838この結果だと微妙
>>714を支持しているように見える
これだと何にも面白くなさそうなんで
>>830のジェファーソンのTは海路はるばる北方へ入植したとしたかったが
>>838この結果だと微妙
842 :世界@名無史さん:2014/01/28(火) 04:31:33.41 0
844 :世界@名無史さん:2014/01/28(火) 16:25:11.05 0
ロシア帝国最後の皇帝、ニコライ2世の5人の子供達
オリガ
http://ja.wikipedia.org/wiki/%E3%82%AA%E3%83%AA%E3%82%AC%E3%83%BB%E3%83%8B%E3%82%B3%E3%83%A9%E3%82%A8%E3%83%B4%E3%83%8A_(%E3%83%8B%E3%82%B3%E3%83%A9%E3%82%A42%E4%B8%96%E7%9A%87%E5%A5%B3)
タチアナ
http://ja.wikipedia.org/wiki/%E3%82%BF%E3%83%81%E3%82%A2%E3%83%8A%E3%83%BB%E3%83%8B%E3%82%B3%E3%83%A9%E3%82%A8%E3%83%B4%E3%83%8A
マリア
http://ja.wikipedia.org/wiki/%E3%83%9E%E3%83%AA%E3%82%A2%E3%83%BB%E3%83%8B%E3%82%B3%E3%83%A9%E3%82%A8%E3%83%B4%E3%83%8A_(%E3%83%8B%E3%82%B3%E3%83%A9%E3%82%A42%E4%B8%96%E7%9A%87%E5%A5%B3)
アナスタシア
http://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%8A%E3%82%B9%E3%82%BF%E3%82%B7%E3%82%A2%E3%83%BB%E3%83%8B%E3%82%B3%E3%83%A9%E3%82%A8%E3%83%B4%E3%83%8A
アレクセイ
http://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%AC%E3%82%AF%E3%82%BB%E3%82%A4%E3%83%BB%E3%83%8B%E3%82%B3%E3%83%A9%E3%82%A8%E3%83%B4%E3%82%A3%E3%83%81_(%E3%83%AD%E3%82%B7%E3%82%A2%E7%9A%87%E5%A4%AA%E5%AD%90)
マリアとタチアナが美人、アレクセイは男前
アナスタシアは少し残念な感じだけど
オリガ
http://ja.wikipedia.org/wiki/%E3%82%AA%E3%83%AA%E3%82%AC%E3%83%BB%E3%83%8B%E3%82%B3%E3%83%A9%E3%82%A8%E3%83%B4%E3%83%8A_(%E3%83%8B%E3%82%B3%E3%83%A9%E3%82%A42%E4%B8%96%E7%9A%87%E5%A5%B3)
タチアナ
http://ja.wikipedia.org/wiki/%E3%82%BF%E3%83%81%E3%82%A2%E3%83%8A%E3%83%BB%E3%83%8B%E3%82%B3%E3%83%A9%E3%82%A8%E3%83%B4%E3%83%8A
マリア
http://ja.wikipedia.org/wiki/%E3%83%9E%E3%83%AA%E3%82%A2%E3%83%BB%E3%83%8B%E3%82%B3%E3%83%A9%E3%82%A8%E3%83%B4%E3%83%8A_(%E3%83%8B%E3%82%B3%E3%83%A9%E3%82%A42%E4%B8%96%E7%9A%87%E5%A5%B3)
アナスタシア
http://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%8A%E3%82%B9%E3%82%BF%E3%82%B7%E3%82%A2%E3%83%BB%E3%83%8B%E3%82%B3%E3%83%A9%E3%82%A8%E3%83%B4%E3%83%8A
アレクセイ
http://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%AC%E3%82%AF%E3%82%BB%E3%82%A4%E3%83%BB%E3%83%8B%E3%82%B3%E3%83%A9%E3%82%A8%E3%83%B4%E3%82%A3%E3%83%81_(%E3%83%AD%E3%82%B7%E3%82%A2%E7%9A%87%E5%A4%AA%E5%AD%90)
マリアとタチアナが美人、アレクセイは男前
アナスタシアは少し残念な感じだけど
ゲノムからのアプローチは方法論的に非常にまずいらしい。
>>829>>838>>841>>846
というかここ>>697にほとんど載ってるね
April 26, 2012
なんだが、、、ワロス
Ancient DNA from Neolithic Sweden (Skoglund et al. 2012)
http://1.bp.blogspot.com/-rl1bw36vh5E/T5WCj36-KyI/AAAAAAAAEyQ/IgbXJAn3FPo/s1600/skoglund.png
http://2.bp.blogspot.com/-JXffzwQQek0/T5WDLwnr2fI/AAAAAAAAEyY/TuDjzH5UJI0/s1600/allele_sharing.png
The Neolithic hunter-gatherers shared most alleles with Northern Europeans, and the lowest allele sharing was with populations from Southeastern Europe (Fig. 3A).
In contrast, the Neolithic farmer shared the greatest fraction of alleles with Southeastern European populations (Cypriots and Greeks),
and showed a pattern of decreasing genetic similarity for populations from the Northwest and Northeast extremes of Europe (Fig. 3B).
Individuals from Turkey stand out by low levels of allele sharing with both Neolithic groups, possibly due to gene flow from outside of Europe
元はこれらしい
Science 27 April 2012: Vol. 336 no. 6080 pp. 466-469 DOI: 10.1126/science.1216304
Origins and Genetic Legacy of Neolithic Farmers and Hunter-Gatherers in Europe
というかここ>>697にほとんど載ってるね
April 26, 2012
なんだが、、、ワロス
Ancient DNA from Neolithic Sweden (Skoglund et al. 2012)
http://1.bp.blogspot.com/-rl1bw36vh5E/T5WCj36-KyI/AAAAAAAAEyQ/IgbXJAn3FPo/s1600/skoglund.png
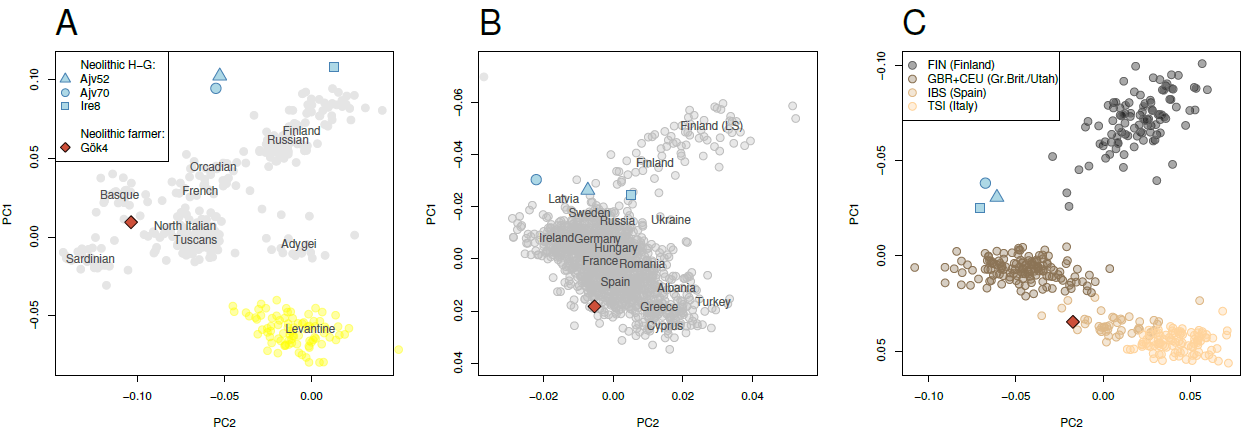{kind=link}
http://2.bp.blogspot.com/-JXffzwQQek0/T5WDLwnr2fI/AAAAAAAAEyY/TuDjzH5UJI0/s1600/allele_sharing.png
{kind=link}
The Neolithic hunter-gatherers shared most alleles with Northern Europeans, and the lowest allele sharing was with populations from Southeastern Europe (Fig. 3A).
In contrast, the Neolithic farmer shared the greatest fraction of alleles with Southeastern European populations (Cypriots and Greeks),
and showed a pattern of decreasing genetic similarity for populations from the Northwest and Northeast extremes of Europe (Fig. 3B).
Individuals from Turkey stand out by low levels of allele sharing with both Neolithic groups, possibly due to gene flow from outside of Europe
元はこれらしい
Science 27 April 2012: Vol. 336 no. 6080 pp. 466-469 DOI: 10.1126/science.1216304
Origins and Genetic Legacy of Neolithic Farmers and Hunter-Gatherers in Europe
>>847
のんびりと検証してきたものがあっさり書いてあってワラタ
まぁ、新しいデータで検証してもほとんど変わらんからいいや
これもあっさり書いてあるけど、>>832
A strong hint for this can also be found in the quite unexpectedly low "TRB" allele sharing of groups from the Northwestern Balkans.
This is quite unexpected, as the area is widely believed to be a conduit through which agriculture spread into Central Europe.
まぁ、これもおいらの結論はあんまり変わらないからいいけどね
のんびりと検証してきたものがあっさり書いてあってワラタ
まぁ、新しいデータで検証してもほとんど変わらんからいいや
これもあっさり書いてあるけど、>>832
A strong hint for this can also be found in the quite unexpectedly low "TRB" allele sharing of groups from the Northwestern Balkans.
This is quite unexpected, as the area is widely believed to be a conduit through which agriculture spread into Central Europe.
まぁ、これもおいらの結論はあんまり変わらないからいいけどね
>>727>>726>>730
これとかもワロス
超古代文明
http://dienekes.blogspot.jp/2014/01/6500-year-old-tin-bronzes-from-serbia.html
6,500-year old tin bronze from Serbia
Antiquity Volume: 87 Number: 338 Page: 1030?1045
Tainted ores and the rise of tin bronzes in Eurasia, c. 6500 years ago
Miljana Radivojevi? et al.
The earliest tin bronze artefacts in Eurasia are generally believed to have appeared in the Near East in the early third millennium BC.
Here we present tin bronze artefacts that occur far from the Near East, and in a significantly earlier period. Excavations at Plo?nik, a Vin?a culture site in Serbia,
recovered a piece of tin bronze foil from an occupation layer dated to the mid fifth millennium BC.
The discovery prompted a reassessment of 14 insufficiently contextualised early tin bronze artefacts from the Balkans.
They too were found to derive from the smelting of copper-tin ores.
These tin bronzes extend the record of bronze making by c. 1500 years, and challenge the conventional narrative of Eurasian metallurgical development.
これとかもワロス
超古代文明
http://dienekes.blogspot.jp/2014/01/6500-year-old-tin-bronzes-from-serbia.html
6,500-year old tin bronze from Serbia
Antiquity Volume: 87 Number: 338 Page: 1030?1045
Tainted ores and the rise of tin bronzes in Eurasia, c. 6500 years ago
Miljana Radivojevi? et al.
The earliest tin bronze artefacts in Eurasia are generally believed to have appeared in the Near East in the early third millennium BC.
Here we present tin bronze artefacts that occur far from the Near East, and in a significantly earlier period. Excavations at Plo?nik, a Vin?a culture site in Serbia,
recovered a piece of tin bronze foil from an occupation layer dated to the mid fifth millennium BC.
The discovery prompted a reassessment of 14 insufficiently contextualised early tin bronze artefacts from the Balkans.
They too were found to derive from the smelting of copper-tin ores.
These tin bronzes extend the record of bronze making by c. 1500 years, and challenge the conventional narrative of Eurasian metallurgical development.
>>682>>683
Ancient Y chromosomes from the Canary Islands
http://dienekes.blogspot.jp/2009/08/ancient-y-chromosomes-from-canary.html
On the aboriginal remains (ABO):
Aboriginal remains were clearly pre-conquest for all the analyzed islands:
Tenerife (2210 ± 60 to 1720 ± 60 BP),
Gomera (1743 ± 40 to 1493 ± 40 BP),
Hierro (1740 ± 50 to 970 ± 50 BP) and
Gran Canaria (1410 ± 60 to 750 ± 60 BP) [33].
It is clear that the aboriginal population was dominated by haplogroups E-M81, E-M78, J-M267.
In the historical period (a few centuries ago) new haplogroups make their appearance (e.g., R1a) and a massive increase in the frequency of R1b is observed.
>>833
Ancient Y chromosomes from the Canary Islands
http://dienekes.blogspot.jp/2009/08/ancient-y-chromosomes-from-canary.html
On the aboriginal remains (ABO):
Aboriginal remains were clearly pre-conquest for all the analyzed islands:
Tenerife (2210 ± 60 to 1720 ± 60 BP),
Gomera (1743 ± 40 to 1493 ± 40 BP),
Hierro (1740 ± 50 to 970 ± 50 BP) and
Gran Canaria (1410 ± 60 to 750 ± 60 BP) [33].
It is clear that the aboriginal population was dominated by haplogroups E-M81, E-M78, J-M267.
In the historical period (a few centuries ago) new haplogroups make their appearance (e.g., R1a) and a massive increase in the frequency of R1b is observed.
>>833
Y-chromosome, mtDNA, and autosomal DNA from Treilles (5,000 years ago, Neolithic France)
http://dienekes.blogspot.jp/2011/05/y-chromosome-mtdna-and-autosomal-dna.html
The paper is behind a paywall, but there is plentiful raw genetic data in the online supplement.
I'll probably have much more to say on this when I read it, but here's the groundbreaking part:
Most of this sample belonged to haplogroup G2a-P15 with some I2a-P37.2 also represented.
>>767?
UPDATE IV:
Table S4 lists (in %) shared mtDNA lineages between Treilles and modern populations.
The top ones are:
Welsh (17.391),
Cornish (16.667),
Central Greeks (14.286),
Bulgarians (12.5).
Several Italian groups as well as South Tyrol Ladins and Germans are also greater than 10%.
http://3.bp.blogspot.com/-a9yFwkju2UU/TeVVmTZ6D8I/AAAAAAAADzw/qWptkbj-evk/s1600/g2a.png(たぶん、、、)
http://dienekes.blogspot.jp/2011/05/y-chromosome-mtdna-and-autosomal-dna.html
The paper is behind a paywall, but there is plentiful raw genetic data in the online supplement.
I'll probably have much more to say on this when I read it, but here's the groundbreaking part:
Most of this sample belonged to haplogroup G2a-P15 with some I2a-P37.2 also represented.
>>767?
UPDATE IV:
Table S4 lists (in %) shared mtDNA lineages between Treilles and modern populations.
The top ones are:
Welsh (17.391),
Cornish (16.667),
Central Greeks (14.286),
Bulgarians (12.5).
Several Italian groups as well as South Tyrol Ladins and Germans are also greater than 10%.
http://3.bp.blogspot.com/-a9yFwkju2UU/TeVVmTZ6D8I/AAAAAAAADzw/qWptkbj-evk/s1600/g2a.png(たぶん、、、)
{kind=link}
>>851
アドレス見たらG2aですね
Welsh
ウェールズ語
http://ja.wikipedia.org/wiki/%E3%82%A6%E3%82%A7%E3%83%BC%E3%83%AB%E3%82%BA%E8%AA%9E
なはっはww
北コーカサスに進入する西地中海人
北中央ヨーロッパ人が中東に侵入
アドレス見たらG2aですね
Welsh
ウェールズ語
http://ja.wikipedia.org/wiki/%E3%82%A6%E3%82%A7%E3%83%BC%E3%83%AB%E3%82%BA%E8%AA%9E
なはっはww
北コーカサスに進入する西地中海人
北中央ヨーロッパ人が中東に侵入
>>851
コーンウォール語
http://en.wikipedia.org/wiki/Cornish_language
7 H*
6 J
4 U5
3 HV0
3 X
2 U5 or K2b1
2 T2b
2 K
1 H
1 I
1 U5b1c
1 H?
1 X2
34 総計
なんか失敗した気がしないでもないけど、まぁいいや
コーンウォール語
http://en.wikipedia.org/wiki/Cornish_language
7 H*
6 J
4 U5
3 HV0
3 X
2 U5 or K2b1
2 T2b
2 K
1 H
1 I
1 U5b1c
1 H?
1 X2
34 総計
なんか失敗した気がしないでもないけど、まぁいいや
854 :世界@名無史さん:2014/01/29(水) 07:58:27.98 0
855 :世界@名無史さん:2014/01/29(水) 09:42:46.63 0
>>847 トルコ人はやはりモンゴロイドの末裔だったか。あるいは gene flow from outside of Europe は中東からか
856 :世界@名無史さん:2014/01/29(水) 12:43:59.07 0
スラブ人は2つの系統があるな
遺伝子から見るとポーランドはゲルマン人、ロシア人はフィン人だね
遺伝子から見るとポーランドはゲルマン人、ロシア人はフィン人だね
859 :世界@名無史さん:2014/01/29(水) 21:50:46.93 0
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=20140127006
↑
日本語でも紹介されている。
青い目と白い肌とはもたらす遺伝子が違うことはわかっているが、
青い目の方が先に欧州人に普及していたようだ。
↑
日本語でも紹介されている。
青い目と白い肌とはもたらす遺伝子が違うことはわかっているが、
青い目の方が先に欧州人に普及していたようだ。
>>745>>838>>841
フィンランド→エストニア、ラトビア
フィンランドの地理
http://ja.wikipedia.org/wiki/%E3%83%95%E3%82%A3%E3%83%B3%E3%83%A9%E3%83%B3%E3%83%89%E3%81%AE%E5%9C%B0%E7%90%86
フィンランドの地理は他の北欧諸国とは違っている。
スウェーデンとの間はバルト海のボスニア湾で境を分かち、ロシアとの間にはフィンランド湾がある。
フィンランドは欧州大陸最北の国である。
フィンランドの地理においてもっとも顕著な傾向は大陸的氷河の痕跡である。
氷河は国土の表面に溝をつくり、磨きあげた。氷河は1万年前に後退したと考えられ、このときモレーン、氷堆丘、エスカーなどを残した。
他にも氷河の存在の兆候として、南部地帯の多数の湖がある。分厚い氷の膜の動く力は、氷床をえぐり、氷が解けるとその穴に水がたまった。
この湖や河川が氷河による形成の痕跡と考えられている。氷河の後退は地学的規模からみればそう遠くない過去のことであり、現代も流域の状況は未完成で、確立したものではない。
氷河の進行と後退の方向はフィンランドの河川や湖の向きを広範囲で北東から南西の向きへ調整した。
サルパウセルッカ分水嶺と呼ばれる二本の峰が25kmの間隔で平行に伸びており、これが氷堆石の終点である。
フィンランド南部にある分水嶺の最高点は標高200mにおよぶ。
3.1 フィンランド多島海域
3.2 フィンランド沿岸部
3.3 湖水地方
3.4 フィンランド高地
フィンランド→エストニア、ラトビア
フィンランドの地理
http://ja.wikipedia.org/wiki/%E3%83%95%E3%82%A3%E3%83%B3%E3%83%A9%E3%83%B3%E3%83%89%E3%81%AE%E5%9C%B0%E7%90%86
フィンランドの地理は他の北欧諸国とは違っている。
スウェーデンとの間はバルト海のボスニア湾で境を分かち、ロシアとの間にはフィンランド湾がある。
フィンランドは欧州大陸最北の国である。
フィンランドの地理においてもっとも顕著な傾向は大陸的氷河の痕跡である。
氷河は国土の表面に溝をつくり、磨きあげた。氷河は1万年前に後退したと考えられ、このときモレーン、氷堆丘、エスカーなどを残した。
他にも氷河の存在の兆候として、南部地帯の多数の湖がある。分厚い氷の膜の動く力は、氷床をえぐり、氷が解けるとその穴に水がたまった。
この湖や河川が氷河による形成の痕跡と考えられている。氷河の後退は地学的規模からみればそう遠くない過去のことであり、現代も流域の状況は未完成で、確立したものではない。
氷河の進行と後退の方向はフィンランドの河川や湖の向きを広範囲で北東から南西の向きへ調整した。
サルパウセルッカ分水嶺と呼ばれる二本の峰が25kmの間隔で平行に伸びており、これが氷堆石の終点である。
フィンランド南部にある分水嶺の最高点は標高200mにおよぶ。
3.1 フィンランド多島海域
3.2 フィンランド沿岸部
3.3 湖水地方
3.4 フィンランド高地
>>861
http://en.wikipedia.org/wiki/Baltic_ice_lake
Eemian Sea, 130,000?115,000 (years ago)
(Baltic ice lake), 12,600?10,300)
Yoldia Sea, 10,300?9500
Ancylus Lake, 9,500?8,000
Mastogloia Sea 8,000?7,500
Littorina Sea, 7,500?4,000
Post-littorina Baltic Sea, 4,000?present
http://en.wikipedia.org/wiki/Baltic_ice_lake
Eemian Sea, 130,000?115,000 (years ago)
(Baltic ice lake), 12,600?10,300)
Yoldia Sea, 10,300?9500
Ancylus Lake, 9,500?8,000
Mastogloia Sea 8,000?7,500
Littorina Sea, 7,500?4,000
Post-littorina Baltic Sea, 4,000?present
>>838
http://fennoscandia.blogspot.jp/2013/08/ajv52-and-european-variation.html
The Ajv52 like Ajv70 and Ire8 individual belonged to a 2800-2000 BC Pitted Ware Culture complex found in Ajvide, Gotland, Sweden.
Ajv52 belonged to mtDNA hg V. The PWC was a hunter gatherer culture.
http://fennoscandia.blogspot.jp/2013/08/ajv52-and-european-variation.html
The Ajv52 like Ajv70 and Ire8 individual belonged to a 2800-2000 BC Pitted Ware Culture complex found in Ajvide, Gotland, Sweden.
Ajv52 belonged to mtDNA hg V. The PWC was a hunter gatherer culture.
寝るタイミングを逸してしまった、、、
>>846>>863
ウラル系の民族の侵入を受けたのが4800年以降で
これらの先住民が居た地域には地中海人が農業を持ち込んだが
フィンランドだけは馴染まなかったという説明だと
シンプルなんだけど、まぁ違和感が残るんだよな
>>846>>863
ウラル系の民族の侵入を受けたのが4800年以降で
これらの先住民が居た地域には地中海人が農業を持ち込んだが
フィンランドだけは馴染まなかったという説明だと
シンプルなんだけど、まぁ違和感が残るんだよな
>>850
航海者ハンノ
http://ja.wikipedia.org/wiki/%E8%88%AA%E6%B5%B7%E8%80%85%E3%83%8F%E3%83%B3%E3%83%8E
航海者ハンノ(こうかいしゃハンノ、英: Hanno the Navigator, Hanno II of Carthage)は、
紀元前450年頃のカルタゴ出身の人物。地中海を越えてアフリカ大陸西岸部にまでの大航海を遂行した。
この功績により後世では他の「ハンノ」たちと区別して『航海者』と名づけられている。
「ハンノ」という意味はフェニキア語で「慈悲深い」という意味である。
航海者ハンノ
http://ja.wikipedia.org/wiki/%E8%88%AA%E6%B5%B7%E8%80%85%E3%83%8F%E3%83%B3%E3%83%8E
航海者ハンノ(こうかいしゃハンノ、英: Hanno the Navigator, Hanno II of Carthage)は、
紀元前450年頃のカルタゴ出身の人物。地中海を越えてアフリカ大陸西岸部にまでの大航海を遂行した。
この功績により後世では他の「ハンノ」たちと区別して『航海者』と名づけられている。
「ハンノ」という意味はフェニキア語で「慈悲深い」という意味である。
867 :世界@名無史さん:2014/01/30(木) 23:09:27.20 0
242 :優しい名無しさん:2014/01/30(木) 10:56:00.88 ID:1NDUICd1
>>230
実際にテレ朝系のビーイングのロックユニット
なんか盗作裁判になってもおかしくないレベルだぜ??w
それが『オマージュ』『リスペクト』。
243 :優しい名無しさん:2014/01/30(木) 10:58:23.33 ID:1NDUICd1
異常な外国文化崇拝。
洋服まで。
こりゃ精神文化的にも珍しい部類じゃないか??
244 :優しい名無しさん:2014/01/30(木) 15:57:02.88 ID:vfvMFGAH
ムーディ勝山のチャラチャッチャラチャーと
桂三度の12 サンっ
でみんな爆笑している時 日本の知性を感じた
良い国だなぁ
>>230
実際にテレ朝系のビーイングのロックユニット
なんか盗作裁判になってもおかしくないレベルだぜ??w
それが『オマージュ』『リスペクト』。
243 :優しい名無しさん:2014/01/30(木) 10:58:23.33 ID:1NDUICd1
異常な外国文化崇拝。
洋服まで。
こりゃ精神文化的にも珍しい部類じゃないか??
244 :優しい名無しさん:2014/01/30(木) 15:57:02.88 ID:vfvMFGAH
ムーディ勝山のチャラチャッチャラチャーと
桂三度の12 サンっ
でみんな爆笑している時 日本の知性を感じた
良い国だなぁ
Lactase persistence
乳糖分解酵素(ラクターゼ)活性持続症
http://www.eps1.comlink.ne.jp/~mayus/eating1-10/lactase.html
なお、馬乳の乳糖濃度は6.3%と高いので、これを発酵させて馬乳酒(アイラグ)を作った。
>>656>>757
http://en.wikipedia.org/wiki/Lactase_persistence
Evolutionary history
Several genetic markers for lactase persistence have been identified,
and these show that the allele has Multiple Origins in Different Parts of the World (i.e. it is an example of convergent evolution).[54]
The version of the allele most common amongst Europeans is estimated to have risen to significant frequencies
about 7,500 years ago in the central Balkans and Central Europe, a place and time approximately corresponding to the archaeological Linearbandkeramik and Star?evo cultures.
From there, it most probably spread eastwards as far as India.
Likewise, one of the four alleles associated with lactase persistence in African population, is also probably of European origin.[55]
Since North Africans also possess this version of the allele it is probable that it actually originated earlier, in the Near East,
but that the earliest farmers did not have high levels of lactase persistence and, subsequently, did not consume significant amounts of unprocessed milk.[56]
Lactase persistence in Sub-Saharan Africa almost certainly had a separate origin, probably more than one,[57]
and it is also likely that there was a separate origin associated with the domestication of the Arabian camel.[58]
None of the mutations so far identified have been shown to be causal for the lactase persistence allele, and it is thought that there are several more yet to be discovered.[59]
乳糖分解酵素(ラクターゼ)活性持続症
http://www.eps1.comlink.ne.jp/~mayus/eating1-10/lactase.html
なお、馬乳の乳糖濃度は6.3%と高いので、これを発酵させて馬乳酒(アイラグ)を作った。
>>656>>757
http://en.wikipedia.org/wiki/Lactase_persistence
Evolutionary history
Several genetic markers for lactase persistence have been identified,
and these show that the allele has Multiple Origins in Different Parts of the World (i.e. it is an example of convergent evolution).[54]
The version of the allele most common amongst Europeans is estimated to have risen to significant frequencies
about 7,500 years ago in the central Balkans and Central Europe, a place and time approximately corresponding to the archaeological Linearbandkeramik and Star?evo cultures.
From there, it most probably spread eastwards as far as India.
Likewise, one of the four alleles associated with lactase persistence in African population, is also probably of European origin.[55]
Since North Africans also possess this version of the allele it is probable that it actually originated earlier, in the Near East,
but that the earliest farmers did not have high levels of lactase persistence and, subsequently, did not consume significant amounts of unprocessed milk.[56]
Lactase persistence in Sub-Saharan Africa almost certainly had a separate origin, probably more than one,[57]
and it is also likely that there was a separate origin associated with the domestication of the Arabian camel.[58]
None of the mutations so far identified have been shown to be causal for the lactase persistence allele, and it is thought that there are several more yet to be discovered.[59]
>>868
Direct estimates of natural selection in Iberia indicate calcium absorption was not the only driver of lactase persistence in Europe
http://mbe.oxfordjournals.org/content/early/2014/01/20/molbev.msu049
Abstract(から抜粋)
In this study we examine one of the most widely cited hypotheses for selection on LP ? that fresh milk consumption supplements the poor vitamin D and calcium status of northern Europe's early farmers (the calcium assimilation hypothesis).
We do this by testing for natural selection on -13,910*T using ancient DNA data from the skeletal remains of eight late Neolithic Iberian individuals,
whom we would not expect to have poor vitamin D and calcium status because of relatively high incident UVB-light levels.
None of the 8 samples successfully typed in the study had the derived T-allele.
In addition, we reanalyse published data from French Neolithic remains to both test for population continuity and further examine the evolution of LP in the region.
Using simulations that accommodate genetic drift, natural selection, uncertainty in calibrated radiocarbon dates, and sampling error,
we find that natural selection is still required to explain the observed increase in allele frequency.
We conclude that the calcium assimilation hypothesis is insufficient to explain the spread of lactase persistence in Europe.
これは結構おもしろいな
the skeletal remains of eight late Neolithic Iberian individuals
French Neolithic remains はたぶん?と思って書き間違いに気が付いた
Direct estimates of natural selection in Iberia indicate calcium absorption was not the only driver of lactase persistence in Europe
http://mbe.oxfordjournals.org/content/early/2014/01/20/molbev.msu049
Abstract(から抜粋)
In this study we examine one of the most widely cited hypotheses for selection on LP ? that fresh milk consumption supplements the poor vitamin D and calcium status of northern Europe's early farmers (the calcium assimilation hypothesis).
We do this by testing for natural selection on -13,910*T using ancient DNA data from the skeletal remains of eight late Neolithic Iberian individuals,
whom we would not expect to have poor vitamin D and calcium status because of relatively high incident UVB-light levels.
None of the 8 samples successfully typed in the study had the derived T-allele.
In addition, we reanalyse published data from French Neolithic remains to both test for population continuity and further examine the evolution of LP in the region.
Using simulations that accommodate genetic drift, natural selection, uncertainty in calibrated radiocarbon dates, and sampling error,
we find that natural selection is still required to explain the observed increase in allele frequency.
We conclude that the calcium assimilation hypothesis is insufficient to explain the spread of lactase persistence in Europe.
これは結構おもしろいな
the skeletal remains of eight late Neolithic Iberian individuals
French Neolithic remains はたぶん?と思って書き間違いに気が付いた
Ancient DNA reveals male diffusion through the Neolithic Mediterranean route
http://www.pnas.org/content/early/2011/05/24/1100723108
Abstract
The Neolithic is a key period in the history of the European settlement.
Although archaeological and present-day genetic data suggest several hypotheses regarding the human migration patterns at this period,
validation of these hypotheses with the use of ancient genetic data has been limited.
In this context, we studied DNA extracted from 53 individuals buried in a necropolis used by a French local community 5,000 y ago.
The relatively good DNA preservation of the samples allowed us to obtain autosomal, Y-chromosomal, and/or mtDNA data for 29 of the 53 samples studied.
From these datasets, we established close parental relationships within the necropolis and determined maternal and paternal lineages
as well as the absence of an allele associated with lactase persistence, probably carried by Neolithic cultures of central Europe.
Our study provides an integrative view of the genetic past in southern France at the end of the Neolithic period.
Furthermore, the Y-haplotype lineages characterized and the study of their current repartition in European populations
confirm a greater influence of the Mediterranean than the Central European route in the peopling of southern Europe during the Neolithic transition.
53体の人骨、29体のY-chromosomal, and/or mtDNA
>>853
>>869たぶんじゃなくてそうだった
http://www.pnas.org/content/early/2011/05/24/1100723108
Abstract
The Neolithic is a key period in the history of the European settlement.
Although archaeological and present-day genetic data suggest several hypotheses regarding the human migration patterns at this period,
validation of these hypotheses with the use of ancient genetic data has been limited.
In this context, we studied DNA extracted from 53 individuals buried in a necropolis used by a French local community 5,000 y ago.
The relatively good DNA preservation of the samples allowed us to obtain autosomal, Y-chromosomal, and/or mtDNA data for 29 of the 53 samples studied.
From these datasets, we established close parental relationships within the necropolis and determined maternal and paternal lineages
as well as the absence of an allele associated with lactase persistence, probably carried by Neolithic cultures of central Europe.
Our study provides an integrative view of the genetic past in southern France at the end of the Neolithic period.
Furthermore, the Y-haplotype lineages characterized and the study of their current repartition in European populations
confirm a greater influence of the Mediterranean than the Central European route in the peopling of southern Europe during the Neolithic transition.
53体の人骨、29体のY-chromosomal, and/or mtDNA
>>853
>>869たぶんじゃなくてそうだった
871 :世界@名無史さん:2014/01/31(金) 22:31:21.77 0
SLC24A5が変化するのは5000〜6000年前ぐらいに始まったということだね。
白色化はまだ続いているのかなあ。
白色化はまだ続いているのかなあ。
>>871
色白好きだなww
>>868
ちなみに7500年前のドイツ農耕民もルクセンブルク狩猟民もラクターゼ活性持続症は持ってない。
だけど現在の北欧では支配的な変異となってる。
しばらく前はアナトリア起源となってたけど、もうちょっと複雑な話になるかもね
色白好きだなww
>>868
ちなみに7500年前のドイツ農耕民もルクセンブルク狩猟民もラクターゼ活性持続症は持ってない。
だけど現在の北欧では支配的な変異となってる。
しばらく前はアナトリア起源となってたけど、もうちょっと複雑な話になるかもね
873 :世界@名無史さん:2014/02/01(土) 03:42:38.52 0
バルカンと中欧起源で、インドのラクターゼ活性持続症と同じタイプらしいな。同じものはアフリカの一部でも見られるらしい
あとはアフリカ起源のが複数と中東のラクダ牧畜民由来のが。インドとヨーロッパの間の地域で見られるものがヨーロッパ起源のものか、中東由来のものかが気になる
ともかく多系統らしい。ヨーロッパ以外のは、牛乳を沸かすための薪が欠乏してやむなく生で飲むうちに...ってのが自然選択のきっかけとして考えられるな。
でも非加熱乳のままでも発酵食品や加工食品は作れるか。
あとはアフリカ起源のが複数と中東のラクダ牧畜民由来のが。インドとヨーロッパの間の地域で見られるものがヨーロッパ起源のものか、中東由来のものかが気になる
ともかく多系統らしい。ヨーロッパ以外のは、牛乳を沸かすための薪が欠乏してやむなく生で飲むうちに...ってのが自然選択のきっかけとして考えられるな。
でも非加熱乳のままでも発酵食品や加工食品は作れるか。
>>656
あとこれがあれば大体が出揃うかな
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=20110708002
イギリスで新石器時代の人骨を大量発見
James Owen for National Geographic News
July 8, 2011
イギリス、スコットランド北部にあるオークニー諸島のサウス・ロナルドセー島で、約5000年前の墓が発見された。内部には新石器時代の大量の人骨が納められていた。
6月に発表されたギブソン氏の報告によると、「バンクス墓(Banks Tomb)」と命名されたこの遺跡は、部分的な発掘調査により、新石器時代にまで遡ると確認された。
荒らされていない新石器時代の墓がスコットランドで見つかったのは約30年ぶりだという。
「これほど保存状態が良い遺跡が見つかるのは珍しい。多数の遺骨が納められており、それぞれ別の年代と考えられる」と同氏は述べている。
墓は、幅0.75メートル、奥行き4メートルの中央の部屋と、周囲の砂岩を切り出して造られた4つの小部屋からなる。
中央の部屋は風雨で傷んだ大きな厚板で蓋をされ、石壁と石柱が支えている。
発掘された頭蓋骨の破片は1000個以上。性別、年齢ともにばらばらで、乳児も含まれていた。
人骨はそれぞれ異なるシルト(細かい泥)の層から出土しており、何世代にもわたって使用されていた可能性があるという。
また最初の調査で、人骨の中から先史時代のカワウソの骨と糞が見つかっており、「カワウソの墓(Tomb of the Otters)」とも呼ばれている。
どうも当時の人々は、それほど頻繁に墓を訪れていなかったらしい。
「墓は密閉されておらず、カワウソが出入りを繰り返していた。つまり、1〜2年おきにしか墓参りや埋葬に来ていなかったと考えられる」とギブソン氏は指摘する。
あとこれがあれば大体が出揃うかな
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=20110708002
イギリスで新石器時代の人骨を大量発見
James Owen for National Geographic News
July 8, 2011
イギリス、スコットランド北部にあるオークニー諸島のサウス・ロナルドセー島で、約5000年前の墓が発見された。内部には新石器時代の大量の人骨が納められていた。
6月に発表されたギブソン氏の報告によると、「バンクス墓(Banks Tomb)」と命名されたこの遺跡は、部分的な発掘調査により、新石器時代にまで遡ると確認された。
荒らされていない新石器時代の墓がスコットランドで見つかったのは約30年ぶりだという。
「これほど保存状態が良い遺跡が見つかるのは珍しい。多数の遺骨が納められており、それぞれ別の年代と考えられる」と同氏は述べている。
墓は、幅0.75メートル、奥行き4メートルの中央の部屋と、周囲の砂岩を切り出して造られた4つの小部屋からなる。
中央の部屋は風雨で傷んだ大きな厚板で蓋をされ、石壁と石柱が支えている。
発掘された頭蓋骨の破片は1000個以上。性別、年齢ともにばらばらで、乳児も含まれていた。
人骨はそれぞれ異なるシルト(細かい泥)の層から出土しており、何世代にもわたって使用されていた可能性があるという。
また最初の調査で、人骨の中から先史時代のカワウソの骨と糞が見つかっており、「カワウソの墓(Tomb of the Otters)」とも呼ばれている。
どうも当時の人々は、それほど頻繁に墓を訪れていなかったらしい。
「墓は密閉されておらず、カワウソが出入りを繰り返していた。つまり、1〜2年おきにしか墓参りや埋葬に来ていなかったと考えられる」とギブソン氏は指摘する。
>>874
オークニー考古学研究センター(Orkney Research Centre for Archaeology)が率いる発掘調査はまだ始まったばかりだが、
「新石器時代の埋葬習慣を解明する重要な手掛かりが得られることは間違いない」とギブソン氏は自信の程をうかがわせる。
例えば、人骨のDNA分析や同位体分析により、血縁関係の有無のほか、
島で緊密な関係を構築していた共同体なのか、外部から来た人間の遺骨も含まれているのかなどを明らかにできるという。
また、儀式的な目的で墓から人骨が取り除かれていないかも調べられる予定だ。
「この墓には非常に多くの遺骨が眠っている。他の石器時代の墓では、これほど多いことは珍しい」。
「カワウソの墓」から1.5キロほど離れた場所には、約5000年前の墓地遺跡「ワシの墓(Tomb of the Eagles)」がある。
最近の調査によると、新石器時代にオークニー諸島で営まれていた酪農集落の生活はあまり平和ではなかったようだ。
ワシの墓から出土した頭蓋骨の20%以上に、鋭利な武器や鈍器で激しく殴打されたとみられる傷跡があった。
カワウソの墓の頭蓋骨にも同じような傷跡がないか、調査が進められている。
「新石器時代の暮らしは、愛と平和をうたうヒッピーのようなイメージがあったが、
実際には暴力的な儀式が行われていた証拠が出てくるかもしれない」とギブソン氏はコメントしている。
オークニー考古学研究センター(Orkney Research Centre for Archaeology)が率いる発掘調査はまだ始まったばかりだが、
「新石器時代の埋葬習慣を解明する重要な手掛かりが得られることは間違いない」とギブソン氏は自信の程をうかがわせる。
例えば、人骨のDNA分析や同位体分析により、血縁関係の有無のほか、
島で緊密な関係を構築していた共同体なのか、外部から来た人間の遺骨も含まれているのかなどを明らかにできるという。
また、儀式的な目的で墓から人骨が取り除かれていないかも調べられる予定だ。
「この墓には非常に多くの遺骨が眠っている。他の石器時代の墓では、これほど多いことは珍しい」。
「カワウソの墓」から1.5キロほど離れた場所には、約5000年前の墓地遺跡「ワシの墓(Tomb of the Eagles)」がある。
最近の調査によると、新石器時代にオークニー諸島で営まれていた酪農集落の生活はあまり平和ではなかったようだ。
ワシの墓から出土した頭蓋骨の20%以上に、鋭利な武器や鈍器で激しく殴打されたとみられる傷跡があった。
カワウソの墓の頭蓋骨にも同じような傷跡がないか、調査が進められている。
「新石器時代の暮らしは、愛と平和をうたうヒッピーのようなイメージがあったが、
実際には暴力的な儀式が行われていた証拠が出てくるかもしれない」とギブソン氏はコメントしている。
>>698
サハラの砂漠化には数千年かかっているという新説
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=18230993
James Owenfor National Geographic News
May 8, 2008
有史以前には緑豊かな草原だったサハラが世界最大の砂漠へと変貌していったペースは、
これまで考えられていたより緩やかだったとする研究報告が発表された。
有史以前の地下水によって維持されているサハラ最大の湖であるヨア湖の湖水には「少なくとも6000年前の気候記録が破壊されずに存在する」と語るのは、
この研究報告の筆者であるケルン大学考古学研究所のシュテファン・クレペリン氏だ。
湖の堆積物から発見された花粉や昆虫、藻などの古代化石は、砂漠化が緩やかに進行したことを示しているという。
この研究報告は、サハラは数百年以内に干上がったとするこれまでの研究を覆すものだ。
従来の研究結果は、大西洋の堆積物から発見された、サハラから風で運ばれたちりに基づくものだった。
「そうした仮説は、サハラで自ら調査したことがない気候学者や科学者が唱えたものであり、
私たちの研究によってサハラの急激な砂漠化は神話だったことがほぼ明らかになった」とクレペリン氏は述べている。
確認されている唯一の急激な変化は、湖そのものが4200年〜3900年前の間に淡水湖から塩湖に変わったことだ。
クレペリン氏によれば、この変化はモンスーンの雨が南へ移動し始めた時期と完全に重なるという。その結果、地表水の流入が止まり、水の蒸発によって塩分が濃くなった。
「この研究報告は、サハラの人口がモンスーンの雨を追って数千年かけて南へ移動したとする従来の考古学上の知見を裏付ける」とクレペリン氏は語る。
サハラの砂漠化には数千年かかっているという新説
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=18230993
James Owenfor National Geographic News
May 8, 2008
有史以前には緑豊かな草原だったサハラが世界最大の砂漠へと変貌していったペースは、
これまで考えられていたより緩やかだったとする研究報告が発表された。
有史以前の地下水によって維持されているサハラ最大の湖であるヨア湖の湖水には「少なくとも6000年前の気候記録が破壊されずに存在する」と語るのは、
この研究報告の筆者であるケルン大学考古学研究所のシュテファン・クレペリン氏だ。
湖の堆積物から発見された花粉や昆虫、藻などの古代化石は、砂漠化が緩やかに進行したことを示しているという。
この研究報告は、サハラは数百年以内に干上がったとするこれまでの研究を覆すものだ。
従来の研究結果は、大西洋の堆積物から発見された、サハラから風で運ばれたちりに基づくものだった。
「そうした仮説は、サハラで自ら調査したことがない気候学者や科学者が唱えたものであり、
私たちの研究によってサハラの急激な砂漠化は神話だったことがほぼ明らかになった」とクレペリン氏は述べている。
確認されている唯一の急激な変化は、湖そのものが4200年〜3900年前の間に淡水湖から塩湖に変わったことだ。
クレペリン氏によれば、この変化はモンスーンの雨が南へ移動し始めた時期と完全に重なるという。その結果、地表水の流入が止まり、水の蒸発によって塩分が濃くなった。
「この研究報告は、サハラの人口がモンスーンの雨を追って数千年かけて南へ移動したとする従来の考古学上の知見を裏付ける」とクレペリン氏は語る。
>>876
また「サハラの気候変動はアフリカ大陸全域、さらには世界の気候変動へとつながる」とクレペリン氏は付け加える。
同氏によれば、ヨア湖での今後の研究から、地球温暖化の進行によるサハラの再緑化の可能性が明らかになるだろうという。
「この可能性に関する確かな情報が得られるものと期待している」とクレペリン氏は語る。
最後の緑地期間が始まったのは1万2000年ほど前で、海洋水の蒸発の増加が原因とみられている。
「これによってもたらされたモンスーンの雨が熱帯地域の地中深く浸透した」という。
「そしていま、人類は同じ状況を生み出そうとしている」とクレペリン氏は語る。
ほぼ30年間サハラ砂漠を研究してきたクレペリン氏によれば、1998年以来、東サハラでは「雨量が増加に転じる強い兆しが見られる」という。
さらに、地域によってはすでに「植生のわずかな変化が観察できる」とも加えた。
あと、こっちも
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=6667009
サハラ砂漠で古代の墓地を発見 よみがえる緑豊かな時代
また「サハラの気候変動はアフリカ大陸全域、さらには世界の気候変動へとつながる」とクレペリン氏は付け加える。
同氏によれば、ヨア湖での今後の研究から、地球温暖化の進行によるサハラの再緑化の可能性が明らかになるだろうという。
「この可能性に関する確かな情報が得られるものと期待している」とクレペリン氏は語る。
最後の緑地期間が始まったのは1万2000年ほど前で、海洋水の蒸発の増加が原因とみられている。
「これによってもたらされたモンスーンの雨が熱帯地域の地中深く浸透した」という。
「そしていま、人類は同じ状況を生み出そうとしている」とクレペリン氏は語る。
ほぼ30年間サハラ砂漠を研究してきたクレペリン氏によれば、1998年以来、東サハラでは「雨量が増加に転じる強い兆しが見られる」という。
さらに、地域によってはすでに「植生のわずかな変化が観察できる」とも加えた。
あと、こっちも
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=6667009
サハラ砂漠で古代の墓地を発見 よみがえる緑豊かな時代
>>656
これは当時は興味なかったけど今では重要なんだろうな
というか、もうDNA解析してる
最古の“核家族”の墓、ドイツで発見
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=2008111802
Carolyn Barryfor National Geographic News
November 17, 2008
母親と父親と2人の男子から成る“核家族”の最古の墓がドイツで発見された。
身を寄せ合うように埋葬されていた4600年前の家族は暴力的な襲撃を受けて亡くなったらしい。
今回の研究から、石器時代後期(紀元前1万年以降)の社会組織を解明する手掛かりも得られそうだ。
この遺骨は2005年にドイツ北東部のユーローの発掘地で複数の墓が集合している場所から出土した。
4つの別々の墓に大人5人、子ども8人の計13人の遺骨が含まれており、
今回のDNA解析の結果から、このグループの中に4〜5歳と8〜9歳の2人の子どもを含む4人家族がいることが分かった。
研究責任者でオーストラリアのアデレード大学のウォルフガング・ハーク氏は、
「このような場所を発見できて運がよかった。通常、家族全員の命が一度に失われることはないし、同時に埋葬されることもない」と話す。
新石器時代に1カ所に複数の墓が作られることはあったが、1人ずつ別々の時期に、場合によっては数年の間隔で埋葬されるのが普通だった。
放射性炭素年代測定で調べた結果、この家族は紀元前2600年頃に生存し、縄目文土器文化に属することが分かった。
縄目文土器(Corded Ware)という名前は当時の土器が特徴的なねじれ模様の圧痕で装飾されていたことに由来している。
ただし、縄目文土器文化の集落が見つかることはめったにないため、考古学者たちは埋葬地から得られる情報に頼ってこの古代文化を解明するしかなかった。
これは当時は興味なかったけど今では重要なんだろうな
というか、もうDNA解析してる
最古の“核家族”の墓、ドイツで発見
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=2008111802
Carolyn Barryfor National Geographic News
November 17, 2008
母親と父親と2人の男子から成る“核家族”の最古の墓がドイツで発見された。
身を寄せ合うように埋葬されていた4600年前の家族は暴力的な襲撃を受けて亡くなったらしい。
今回の研究から、石器時代後期(紀元前1万年以降)の社会組織を解明する手掛かりも得られそうだ。
この遺骨は2005年にドイツ北東部のユーローの発掘地で複数の墓が集合している場所から出土した。
4つの別々の墓に大人5人、子ども8人の計13人の遺骨が含まれており、
今回のDNA解析の結果から、このグループの中に4〜5歳と8〜9歳の2人の子どもを含む4人家族がいることが分かった。
研究責任者でオーストラリアのアデレード大学のウォルフガング・ハーク氏は、
「このような場所を発見できて運がよかった。通常、家族全員の命が一度に失われることはないし、同時に埋葬されることもない」と話す。
新石器時代に1カ所に複数の墓が作られることはあったが、1人ずつ別々の時期に、場合によっては数年の間隔で埋葬されるのが普通だった。
放射性炭素年代測定で調べた結果、この家族は紀元前2600年頃に生存し、縄目文土器文化に属することが分かった。
縄目文土器(Corded Ware)という名前は当時の土器が特徴的なねじれ模様の圧痕で装飾されていたことに由来している。
ただし、縄目文土器文化の集落が見つかることはめったにないため、考古学者たちは埋葬地から得られる情報に頼ってこの古代文化を解明するしかなかった。
>>878
今回の新発見は、当時の社会で家族関係が重要な役割を担っていたことを示唆するものだ。
大部分の遺骨は男性たちが一方向を向き、女性たちが別の一方向を向いて埋葬されていたが、
この家族の墓では2人の男子が両親の方を向いて埋葬されており、珍しい配置だという。
「論理的な関係を表すことで、この人たちが家族だったことを示したかったのだろう」とハーク氏は言う。
近くの別の墓には男子と女子の遺骨があったが、一緒に埋葬されている大人の女性の方を向いてはいなかった。
遺伝子検査の結果、子どもたちの両親は同じ人だったが、大人の女性は母親ではないことが分かった。ハーク氏は、「彼女は叔母さんか継母だったのかもしれない」と話す。
ロンドン自然史博物館の古生物学者ルイーズ・ハンフリー氏は、「今回の研究は小さな核家族が同時に埋葬されていたことを裏付ける決定的な遺伝子的証拠を提示している。
家族の絆が強く、死に際しても尊重されていたことが分かる」と述べている。
遺骨には新石器時代によく見られる残虐行為の痕跡があった。
ハーク氏によると、ある大人の遺骨の脊椎には矢が刺さっており、頭蓋骨の割れ目や防御的な負傷の跡は「突然の暴力的な最後」を示しているという。
放射性炭素年代測定の結果によると、墓地の埋葬はほぼ同時期に行われ、中には同じ日に埋葬された遺骨もあったようだ。
つまり、この村は襲撃を受けた可能性があり、遺骨を調べたところでは、30歳以上の何人かの大人と10歳未満の子どもたちが襲撃中に殺されたと推測できる。
強靭なメンバーは襲撃を生き延びたか、あるいは狩猟のために外出していたとハーク氏は推測している。
「何が起こったにせよ、生存者が戻ってきて、死者を手厚く埋葬したことは確かだ。だれかが事実を知っていて、それを守り伝えようとしたのだろう」。
今回の新発見は、当時の社会で家族関係が重要な役割を担っていたことを示唆するものだ。
大部分の遺骨は男性たちが一方向を向き、女性たちが別の一方向を向いて埋葬されていたが、
この家族の墓では2人の男子が両親の方を向いて埋葬されており、珍しい配置だという。
「論理的な関係を表すことで、この人たちが家族だったことを示したかったのだろう」とハーク氏は言う。
近くの別の墓には男子と女子の遺骨があったが、一緒に埋葬されている大人の女性の方を向いてはいなかった。
遺伝子検査の結果、子どもたちの両親は同じ人だったが、大人の女性は母親ではないことが分かった。ハーク氏は、「彼女は叔母さんか継母だったのかもしれない」と話す。
ロンドン自然史博物館の古生物学者ルイーズ・ハンフリー氏は、「今回の研究は小さな核家族が同時に埋葬されていたことを裏付ける決定的な遺伝子的証拠を提示している。
家族の絆が強く、死に際しても尊重されていたことが分かる」と述べている。
遺骨には新石器時代によく見られる残虐行為の痕跡があった。
ハーク氏によると、ある大人の遺骨の脊椎には矢が刺さっており、頭蓋骨の割れ目や防御的な負傷の跡は「突然の暴力的な最後」を示しているという。
放射性炭素年代測定の結果によると、墓地の埋葬はほぼ同時期に行われ、中には同じ日に埋葬された遺骨もあったようだ。
つまり、この村は襲撃を受けた可能性があり、遺骨を調べたところでは、30歳以上の何人かの大人と10歳未満の子どもたちが襲撃中に殺されたと推測できる。
強靭なメンバーは襲撃を生き延びたか、あるいは狩猟のために外出していたとハーク氏は推測している。
「何が起こったにせよ、生存者が戻ってきて、死者を手厚く埋葬したことは確かだ。だれかが事実を知っていて、それを守り伝えようとしたのだろう」。
どうもこのスレはたった一人の方が自作自演しているように見えますが、
横から鼻を突っ込ませてもらいますと、縄目紋土器文化の時代は、
のちのゲルマン・ケルト・スラブ・バルト・ラテンがまだ同じひとつの集団だった時代ですね。
この後の文化区分の時代にゲルマン・スラブ・バルトの元になる集団から、
ケルトやラテンの元になる集団が分離していきます。
横から鼻を突っ込ませてもらいますと、縄目紋土器文化の時代は、
のちのゲルマン・ケルト・スラブ・バルト・ラテンがまだ同じひとつの集団だった時代ですね。
この後の文化区分の時代にゲルマン・スラブ・バルトの元になる集団から、
ケルトやラテンの元になる集団が分離していきます。
881 :世界@名無史さん:2014/02/01(土) 13:11:18.90 0
いろいろいなことが個別にわかってくるね。
これを統合すると欧州古代史の全体像がはっきりする。
もう少しか。
これを統合すると欧州古代史の全体像がはっきりする。
もう少しか。
>>849>>858
>>730>>739
Chronology[edit]
The origins of the Vin?a culture are debated.
Before the advent of radiocarbon dating it was thought, on the basis of typological similarities,
that Vin?a and other Neolithic cultures belonging to the 'Dark Burnished Ware' complex were the product of migrations from Anatolia to the Balkans.
This had to be reassessed in light of radiocarbon dates which showed that the Dark Burnished Ware complex appeared at least a millennium before Troy I,
the putative starting point of the westward migration.
An alternative hypothesis where the Vin?a culture developed locally from the preceding Star?evo culture?first proposed by Colin Renfrew in 1969?
is now accepted by many scholars, but the evidence is not conclusive.[3][4]
The Vin?a culture can be divided into two phases, closely linked with those of its type site Vin?a-Belo Brdo:[5]
>>730>>739
Chronology[edit]
The origins of the Vin?a culture are debated.
Before the advent of radiocarbon dating it was thought, on the basis of typological similarities,
that Vin?a and other Neolithic cultures belonging to the 'Dark Burnished Ware' complex were the product of migrations from Anatolia to the Balkans.
This had to be reassessed in light of radiocarbon dates which showed that the Dark Burnished Ware complex appeared at least a millennium before Troy I,
the putative starting point of the westward migration.
An alternative hypothesis where the Vin?a culture developed locally from the preceding Star?evo culture?first proposed by Colin Renfrew in 1969?
is now accepted by many scholars, but the evidence is not conclusive.[3][4]
The Vin?a culture can be divided into two phases, closely linked with those of its type site Vin?a-Belo Brdo:[5]
>>882
>>822
Star?evo culture
http://en.wikipedia.org/wiki/Star%C4%8Devo_culture
The Star?evo culture, sometimes included within a larger grouping known as the Star?evo?K?ros?Cri? culture,[1]
is an archaeological culture of Southeastern Europe, dating to the Neolithic period between c. 5500 and 4500 BCE[2]
(according to other source, between 6200 and 5200 BCE).[3]
Star?evo, the type site, is located on the north bank of the Danube in Serbia (Vojvodina province), opposite Belgrade.
It represents the earliest settled farming society in the area, although hunting and gathering still provided a significant portion of the inhabitants' diet.
http://upload.wikimedia.org/wikipedia/commons/thumb/8/85/Neol%C3%ADtico_en_Europa.png/800px-Neol%C3%ADtico_en_Europa.png
>>822
Star?evo culture
http://en.wikipedia.org/wiki/Star%C4%8Devo_culture
The Star?evo culture, sometimes included within a larger grouping known as the Star?evo?K?ros?Cri? culture,[1]
is an archaeological culture of Southeastern Europe, dating to the Neolithic period between c. 5500 and 4500 BCE[2]
(according to other source, between 6200 and 5200 BCE).[3]
Star?evo, the type site, is located on the north bank of the Danube in Serbia (Vojvodina province), opposite Belgrade.
It represents the earliest settled farming society in the area, although hunting and gathering still provided a significant portion of the inhabitants' diet.
http://upload.wikimedia.org/wikipedia/commons/thumb/8/85/Neol%C3%ADtico_en_Europa.png/800px-Neol%C3%ADtico_en_Europa.png
{kind=link}
>>880
このレスも含めて全部おいらの自演
このレスも含めて全部おいらの自演
885 :世界@名無史さん:2014/02/02(日) 08:04:17.66 0
>>880
分化的統一体であることと、血統的に現在の民族のルーツであることは別だからね。
分化的統一体であることと、血統的に現在の民族のルーツであることは別だからね。
>>885
まぁ、そういうことっすね
>>878
Y-chromosomes and mtDNA from Eulau (Corded Ware Culture, Germany)
http://dienekes.blogspot.jp/2008/11/y-chromosomes-and-mtdna-from-eulau.html
Three male individuals, a 40-60 year-old male and two young boys (4-5 and 8-9 years old) from grave 99 belonged to Y-haplogroup R1a. The boys were related to their parent:
Additionally, the Y chromosome haplogroup R1a of the boys corresponds with the man’s (ind. 3).
The mtDNA results included haplogroups:
K1b (three from grave 99) 16093C, 16224C, 16311C, 16319A
U5b (one from grave 99) 16189C, 16192T, 16270T
I (one from grave 90) 16129A, 16223T, 16391A
H (one from grave 98) 16093C, 16221T
X2 (two from grave 98) 16189C, 16223T, 16278T 73G, 153G, 195C, 225A, 226C, 263G
K1a2 (one from grave 93) 16145A, 16224C, 16311C
たぶんこれ
Ancient DNA, Strontium isotopes, and osteological analyses shed light on social and kinship organization of the Later Stone Age
http://www.pnas.org/content/105/47/18226.short
まぁ、そういうことっすね
>>878
Y-chromosomes and mtDNA from Eulau (Corded Ware Culture, Germany)
http://dienekes.blogspot.jp/2008/11/y-chromosomes-and-mtdna-from-eulau.html
Three male individuals, a 40-60 year-old male and two young boys (4-5 and 8-9 years old) from grave 99 belonged to Y-haplogroup R1a. The boys were related to their parent:
Additionally, the Y chromosome haplogroup R1a of the boys corresponds with the man’s (ind. 3).
The mtDNA results included haplogroups:
K1b (three from grave 99) 16093C, 16224C, 16311C, 16319A
U5b (one from grave 99) 16189C, 16192T, 16270T
I (one from grave 90) 16129A, 16223T, 16391A
H (one from grave 98) 16093C, 16221T
X2 (two from grave 98) 16189C, 16223T, 16278T 73G, 153G, 195C, 225A, 226C, 263G
K1a2 (one from grave 93) 16145A, 16224C, 16311C
たぶんこれ
Ancient DNA, Strontium isotopes, and osteological analyses shed light on social and kinship organization of the Later Stone Age
http://www.pnas.org/content/105/47/18226.short
>>886
気になる部分なので残り全部もコピペ
The occurrence of X2 and K1b suggests changes in frequency to the present-day:
The detection of mtDNA haplotypes X2 and K1b, both being very rare in modern-day European populations,
is unlikely to be based on the occurrence of independent contamination events.
For the X2 sequence:
At present, three exact matches were found among individuals from Iran, Syria, and Estonia showing HVS I and HVS II patterns identical with the two Eulau individuals,
but differing from each other by further coding region polymorphisms.
Regarding the present-day distribution of X2:
Overall, it appears that the populations of the Near East, the Caucasus, and Mediterranean Europe harbor subhaplogroup X2
at higher frequencies than those of northern and northeastern Europe (P less than .05) and that X2 is rare in Eastern European
as well as Central Asian, Siberian, and Indian populations and is virtually absent in the Finno-Ugric and Turkic-speaking people of the Volga-Ural region.
For the K1b sequences:
Today, haplogroup K has a frequency of around 6% in Europe (6, 39).
The identical sequence haplotype of individuals 1, 2, and 4 has previously been observed (4, 5).
...
So far this haplotype has uniquely been reported in two modern Shugnans of Tadzhikistan (5)
気になる部分なので残り全部もコピペ
The occurrence of X2 and K1b suggests changes in frequency to the present-day:
The detection of mtDNA haplotypes X2 and K1b, both being very rare in modern-day European populations,
is unlikely to be based on the occurrence of independent contamination events.
For the X2 sequence:
At present, three exact matches were found among individuals from Iran, Syria, and Estonia showing HVS I and HVS II patterns identical with the two Eulau individuals,
but differing from each other by further coding region polymorphisms.
Regarding the present-day distribution of X2:
Overall, it appears that the populations of the Near East, the Caucasus, and Mediterranean Europe harbor subhaplogroup X2
at higher frequencies than those of northern and northeastern Europe (P less than .05) and that X2 is rare in Eastern European
as well as Central Asian, Siberian, and Indian populations and is virtually absent in the Finno-Ugric and Turkic-speaking people of the Volga-Ural region.
For the K1b sequences:
Today, haplogroup K has a frequency of around 6% in Europe (6, 39).
The identical sequence haplotype of individuals 1, 2, and 4 has previously been observed (4, 5).
...
So far this haplotype has uniquely been reported in two modern Shugnans of Tadzhikistan (5)
>>887
The haplogroup I is also interesting, since it was found at a high frequency in Vikings and Iron Age Danes but has a lower frequency in modern times.
So, in general, these results once again point towards a change in the mtDNA gene pool across the millennia, explained by the authors:
However, we are aware of the fact that the modern distribution might not reflect the haplotype distribution during the Late Neolithic.
The mitochondrial haplogroup distribution of a population is likely to have changed throughout time by factors like genetic drift or events like migration or genetic palimpsest (40).
As we recently showed, the frequencies of haplogroups could have been significantly different between Neolithic and present populations (41),
although a similar study (from a different region in Europe) provides evidence of genetic continuity throughout the millennia (42).
Study (41) also by Haak et al. was about Linearbandkeramik farmers from Central Europe. Study (42) by Sampietro et al. was about Neolithic Iberians.
The haplogroup I is also interesting, since it was found at a high frequency in Vikings and Iron Age Danes but has a lower frequency in modern times.
So, in general, these results once again point towards a change in the mtDNA gene pool across the millennia, explained by the authors:
However, we are aware of the fact that the modern distribution might not reflect the haplotype distribution during the Late Neolithic.
The mitochondrial haplogroup distribution of a population is likely to have changed throughout time by factors like genetic drift or events like migration or genetic palimpsest (40).
As we recently showed, the frequencies of haplogroups could have been significantly different between Neolithic and present populations (41),
although a similar study (from a different region in Europe) provides evidence of genetic continuity throughout the millennia (42).
Study (41) also by Haak et al. was about Linearbandkeramik farmers from Central Europe. Study (42) by Sampietro et al. was about Neolithic Iberians.
890 :世界@名無史さん:2014/02/02(日) 14:59:31.76 0
ここ1〜2年いろいろな研究成果が発表されてはっきりしてきたね。
ここ5000〜6000年内にかなりの変化があったということ。
取って代わったのが東方からの酪農をもたらした農耕民であること。
(騎馬民族とか遊牧民族とはちょっと異なる)
青い目はその前に普及し始めていたこと。
白い肌はたぶん東方からの侵入者がもたらしたということ。
当然印欧語も東方からの侵入者がもたらしたということ。
もう少し私見を入れると、R1bの欧州への侵入は数次にわたるものであり、
欧州へ侵入した印欧語族はR1bの後からの侵入者+R1aということかな。
ここ5000〜6000年内にかなりの変化があったということ。
取って代わったのが東方からの酪農をもたらした農耕民であること。
(騎馬民族とか遊牧民族とはちょっと異なる)
青い目はその前に普及し始めていたこと。
白い肌はたぶん東方からの侵入者がもたらしたということ。
当然印欧語も東方からの侵入者がもたらしたということ。
もう少し私見を入れると、R1bの欧州への侵入は数次にわたるものであり、
欧州へ侵入した印欧語族はR1bの後からの侵入者+R1aということかな。
Iを金髪碧眼薄肌の白人だとする先入観がそうさせている。
実はIは黒人に近く浅黒い、レスピューグのビーナスの作者のハプロタイプ。
金髪碧眼薄肌はハプロタイプR1で決定。
実はIは黒人に近く浅黒い、レスピューグのビーナスの作者のハプロタイプ。
金髪碧眼薄肌はハプロタイプR1で決定。
>>889
これも当時まったく興味なかったやつだね
デンマークで鉄器時代の墓地遺跡から「アラブ系DNA」を確認
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=14651408
James Owenfor National Geographic News
June 24, 2008
デンマークで発掘された古代デーン人のDNAからアラビア系の遺伝子が発見された。
2000年前のスカンジナビア地方には、いまより遺伝子的に多様な人々が生活していたと考えられる。
このデーン人は鉄器時代の男性で、ローマ帝国の北の国境付近を守る兵士、
または中東から連れてこられた女性奴隷の子孫であった可能性がある。
この遺体は、デンマーク東部のシェラン島の南側にあるベゲブヤゴーデという鉄器時代の遺跡で見つかったものだ。
「この遺跡から発掘された遺体は貧しい農民階級に属していた可能性もある」とコペンハーゲン大学法医学研究所のリネア・ネルキオール氏が率いる研究チームは報告している。
また、「今回の分析ではほかにも珍しいハプログループ(遺伝的集団)が発見され、
中には現在デンマークに住む人々にはわずか2%しか見られない有史以前の欧州系統の特徴を示すものもあった。
後にスカンジナビア半島やドイツからの移民によって特徴が薄められた古代北欧人の一種とも考えられる」と同氏は説明する。
一方、デンマーク東部の墓地遺跡で発掘された遺体には、現在、スカンジナビア地方に暮らす人々に共通して見られる遺伝的特徴が備わっていたという。
「こちらの墓地遺跡で見つかった遺体は、主に北欧人型の1種類の遺伝的特徴でまとまっており、多様性はあまり見られない。
彼らは埋葬される前に美しい宝石で装飾されているため、この付近には上流階級の社会が存在したと考えられる。
おそらくはスカンジナビア地方の別の場所からやってきた人々だろう」と同氏は述べている。
研究の結果、古代のデンマークには現在よりもかなり遺伝的に多様な人々が暮らしていたことが示唆された。
これも当時まったく興味なかったやつだね
デンマークで鉄器時代の墓地遺跡から「アラブ系DNA」を確認
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=14651408
James Owenfor National Geographic News
June 24, 2008
デンマークで発掘された古代デーン人のDNAからアラビア系の遺伝子が発見された。
2000年前のスカンジナビア地方には、いまより遺伝子的に多様な人々が生活していたと考えられる。
このデーン人は鉄器時代の男性で、ローマ帝国の北の国境付近を守る兵士、
または中東から連れてこられた女性奴隷の子孫であった可能性がある。
この遺体は、デンマーク東部のシェラン島の南側にあるベゲブヤゴーデという鉄器時代の遺跡で見つかったものだ。
「この遺跡から発掘された遺体は貧しい農民階級に属していた可能性もある」とコペンハーゲン大学法医学研究所のリネア・ネルキオール氏が率いる研究チームは報告している。
また、「今回の分析ではほかにも珍しいハプログループ(遺伝的集団)が発見され、
中には現在デンマークに住む人々にはわずか2%しか見られない有史以前の欧州系統の特徴を示すものもあった。
後にスカンジナビア半島やドイツからの移民によって特徴が薄められた古代北欧人の一種とも考えられる」と同氏は説明する。
一方、デンマーク東部の墓地遺跡で発掘された遺体には、現在、スカンジナビア地方に暮らす人々に共通して見られる遺伝的特徴が備わっていたという。
「こちらの墓地遺跡で見つかった遺体は、主に北欧人型の1種類の遺伝的特徴でまとまっており、多様性はあまり見られない。
彼らは埋葬される前に美しい宝石で装飾されているため、この付近には上流階級の社会が存在したと考えられる。
おそらくはスカンジナビア地方の別の場所からやってきた人々だろう」と同氏は述べている。
研究の結果、古代のデンマークには現在よりもかなり遺伝的に多様な人々が暮らしていたことが示唆された。
>>892
ネルキオール氏の説明によると「信頼できるDNA分析結果としては、これまでに石器時代後期から中世にわたる56人分の結果が得られている」という。
「私たちが調査したデンマークの墳墓すべてで、いまのヨーロッパでは見られない珍しい遺伝子タイプを発見している。
それにも関わらず、デーン人の子孫にあたる人々の遺伝子に多様性があまり見られないのは不思議だ」と同氏は分析する。
この理由として、同研究チームでは、中世に黒死病として名高い腺ペストが大流行したときに、ほかの人々よりもこの病気にかかりやすいグループが存在したのではないかと推測している。
1347〜51年の大流行は欧州の人口の約3分の1の人々を死に追いやっている。
また、中世のイングランドでも同様に、ある特定のハプログループが広がり、遺伝的多様性が狭まったことも分かっている。
「これは、ある遺伝子を持つ家系がペストの大流行から生き残ったという事実を裏付けるものかもしれない」と専門家は話している。
ネルキオール氏の説明によると「信頼できるDNA分析結果としては、これまでに石器時代後期から中世にわたる56人分の結果が得られている」という。
「私たちが調査したデンマークの墳墓すべてで、いまのヨーロッパでは見られない珍しい遺伝子タイプを発見している。
それにも関わらず、デーン人の子孫にあたる人々の遺伝子に多様性があまり見られないのは不思議だ」と同氏は分析する。
この理由として、同研究チームでは、中世に黒死病として名高い腺ペストが大流行したときに、ほかの人々よりもこの病気にかかりやすいグループが存在したのではないかと推測している。
1347〜51年の大流行は欧州の人口の約3分の1の人々を死に追いやっている。
また、中世のイングランドでも同様に、ある特定のハプログループが広がり、遺伝的多様性が狭まったことも分かっている。
「これは、ある遺伝子を持つ家系がペストの大流行から生き残ったという事実を裏付けるものかもしれない」と専門家は話している。
894 :世界@名無史さん:2014/02/02(日) 19:04:48.26 0
>>892
シェラン島
http://ja.wikipedia.org/wiki/%E3%82%B7%E3%82%A7%E3%83%A9%E3%83%B3%E5%B3%B6
シェラン島(デンマーク語:Sjalland)は、デンマークの島。ユトランド半島の東、バルト海に位置する。
面積は7,031 km2でグリーンランドとその属島を除くとデンマークで最も広い。
人口は約210万人。デンマーク王国の東部にあり、東端には首都コペンハーゲンがある。
西にあるフュン島との間には大ベルト海峡を跨ぎ大ベルト橋が架かる。
スウェーデンとの間はエーレスンド海峡を跨ぐエーレスンド橋で結ばれている。
ファルスター島との間はファローブローン橋(デンマーク語版)で結ばれている。
メーラレン湖 - 北欧神話においてシェラン島は女神ゲフィオンがスウェーデンの土地を削り取って作ったと言われ、その跡がこの湖だとされている。
http://ja.wikipedia.org/wiki/%E3%83%A1%E3%83%BC%E3%83%A9%E3%83%AC%E3%83%B3%E6%B9%96
神話[編集]
北欧神話ではメーラレン湖は女神ゲフィオン(Gefjun)がスウェーデンの王ギュルヴィ(Gylfi)をだまして土地を引いた跡地であるとされている[3]。
これは別名ゲフィオンの国引きと言われているが、デンマークでは湖と島の形の相似から首都のあるシェラン島がメーラレン湖から引かれて来た土地と考え、
この伝承を国作り神話と捉えている[4]。
>>862
シェラン島
http://ja.wikipedia.org/wiki/%E3%82%B7%E3%82%A7%E3%83%A9%E3%83%B3%E5%B3%B6
シェラン島(デンマーク語:Sjalland)は、デンマークの島。ユトランド半島の東、バルト海に位置する。
面積は7,031 km2でグリーンランドとその属島を除くとデンマークで最も広い。
人口は約210万人。デンマーク王国の東部にあり、東端には首都コペンハーゲンがある。
西にあるフュン島との間には大ベルト海峡を跨ぎ大ベルト橋が架かる。
スウェーデンとの間はエーレスンド海峡を跨ぐエーレスンド橋で結ばれている。
ファルスター島との間はファローブローン橋(デンマーク語版)で結ばれている。
メーラレン湖 - 北欧神話においてシェラン島は女神ゲフィオンがスウェーデンの土地を削り取って作ったと言われ、その跡がこの湖だとされている。
http://ja.wikipedia.org/wiki/%E3%83%A1%E3%83%BC%E3%83%A9%E3%83%AC%E3%83%B3%E6%B9%96
神話[編集]
北欧神話ではメーラレン湖は女神ゲフィオン(Gefjun)がスウェーデンの王ギュルヴィ(Gylfi)をだまして土地を引いた跡地であるとされている[3]。
これは別名ゲフィオンの国引きと言われているが、デンマークでは湖と島の形の相似から首都のあるシェラン島がメーラレン湖から引かれて来た土地と考え、
この伝承を国作り神話と捉えている[4]。
>>862
>>892
コメントの無さが凄い
Ancient mtDNA from Iron Age Denmark
http://dienekes.blogspot.jp/2007/12/ancient-mtdna-from-iron-age-denmark.html
The Roman Iron-Age (0-400 AD) in Southern Scandinavia was a formative period, where the society changed from archaic chiefdoms to a true state formation,
and the population composition has likely changed in this period due to immigrants from Middle Scandinavia.
We have analyzed mtDNA from 22 individuals from two different types of settlements, Bogebjerggard and Skovgaarde, in Southern Denmark.
Bogebjerggard (ca. 0 AD) represents the lowest level of free, but poor farmers, whereas Skovgaarde 8 km to the east (ca. 200-270 AD) represents the highest level of the society.
Reproducible results were obtained for 18 subjects harboring 17 different haplotypes all compatible (in their character states) with the phylogenetic tree drawn from present day populations of Europe.
This indicates that the South Scandinavian Roman Iron-Age population was as diverse as Europeans are today.
Several of the haplogroups (R0a, U2, I) observed in Bogebjerggard are rare in present day Scandinavians.
Most significantly, R0a, harbored by a male, is a haplogroup frequent in East Africa and Arabia but virtually absent among modern Northern Europeans.
We suggest that this subject was a soldier or a slave, or a descendant of a female slave, from Roman Legions stationed a few hundred kilometers to the south.
In contrast, the haplotype distribution in the rich Skovgaarde shows similarity to that observed for modern Scandinavians, and the Bogebjerggard and Skovgaarde population samples differ significantly (P approximately 0.01).
Skovgaarde may represent a new upper-class formed by migrants from Middle Scandinavia bringing with them Scandinavian haplogroups.
>>892
N1aのことかと思った
コメントの無さが凄い
Ancient mtDNA from Iron Age Denmark
http://dienekes.blogspot.jp/2007/12/ancient-mtdna-from-iron-age-denmark.html
The Roman Iron-Age (0-400 AD) in Southern Scandinavia was a formative period, where the society changed from archaic chiefdoms to a true state formation,
and the population composition has likely changed in this period due to immigrants from Middle Scandinavia.
We have analyzed mtDNA from 22 individuals from two different types of settlements, Bogebjerggard and Skovgaarde, in Southern Denmark.
Bogebjerggard (ca. 0 AD) represents the lowest level of free, but poor farmers, whereas Skovgaarde 8 km to the east (ca. 200-270 AD) represents the highest level of the society.
Reproducible results were obtained for 18 subjects harboring 17 different haplotypes all compatible (in their character states) with the phylogenetic tree drawn from present day populations of Europe.
This indicates that the South Scandinavian Roman Iron-Age population was as diverse as Europeans are today.
Several of the haplogroups (R0a, U2, I) observed in Bogebjerggard are rare in present day Scandinavians.
Most significantly, R0a, harbored by a male, is a haplogroup frequent in East Africa and Arabia but virtually absent among modern Northern Europeans.
We suggest that this subject was a soldier or a slave, or a descendant of a female slave, from Roman Legions stationed a few hundred kilometers to the south.
In contrast, the haplotype distribution in the rich Skovgaarde shows similarity to that observed for modern Scandinavians, and the Bogebjerggard and Skovgaarde population samples differ significantly (P approximately 0.01).
Skovgaarde may represent a new upper-class formed by migrants from Middle Scandinavia bringing with them Scandinavian haplogroups.
>>892
N1aのことかと思った
>>896
これ意外とわかんないね
http://anthropologynet.files.wordpress.com/2008/06/bogebjerggard-skovgaarde-denmark-map.gif
http://www.ncbi.nlm.nih.gov/pubmed?tmpl=NoSidebarfile&cmd=Retrieve&list_uids=18046774&dopt=AbstractPlus
やる気がでたらみる
http://dienekes.blogspot.jp/2010/07/nordicdb-paper-on-finns-danes-and.html
http://dienekes.blogspot.jp/2008/05/ancient-viking-mtdna-from-denmark.html
http://2.bp.blogspot.com/_Ish7688voT0/TFFaawV3n0I/AAAAAAAAChI/GulnWQazmV8/s1600/leunordicdb.jpg
これは?>>616
European hunter-gatherers acquired pigs with Near Eastern and European mtDNA
http://dienekes.blogspot.jp/2013/08/european-hunter-gatherers-acquired-pigs.html
これ意外とわかんないね
http://anthropologynet.files.wordpress.com/2008/06/bogebjerggard-skovgaarde-denmark-map.gif
{kind=link}
http://www.ncbi.nlm.nih.gov/pubmed?tmpl=NoSidebarfile&cmd=Retrieve&list_uids=18046774&dopt=AbstractPlus
やる気がでたらみる
http://dienekes.blogspot.jp/2010/07/nordicdb-paper-on-finns-danes-and.html
http://dienekes.blogspot.jp/2008/05/ancient-viking-mtdna-from-denmark.html
http://2.bp.blogspot.com/_Ish7688voT0/TFFaawV3n0I/AAAAAAAAChI/GulnWQazmV8/s1600/leunordicdb.jpg
{kind=link}
これは?>>616
European hunter-gatherers acquired pigs with Near Eastern and European mtDNA
http://dienekes.blogspot.jp/2013/08/european-hunter-gatherers-acquired-pigs.html
>>897
http://1.bp.blogspot.com/-_mSQwcwmmik/UhzHzuryKTI/AAAAAAAAJE0/Po072BENEqg/s1600/ncomms3348-f1.jpg
This is a nice demonstration of transfer of domesticated animals from Neolithic farmers to European hunter-gatherers. (Red=European, Yellow=Near Eastern).
The comings and goings of Near Eastern and European domestic pigs (Ottoni et al. 2012)
http://dienekes.blogspot.jp/2012/11/the-comings-and-goings-of-near-eastern.html
http://2.bp.blogspot.com/-yQYdErp2j2k/UK8tdYlo0OI/AAAAAAAAHmE/IchI7xSYLNQ/s1600/ottoni.png
The left-right arrangement of the columns corresponds to a west-east longitude across West Asia.
It can be easily seen that some of the early domestic samples (yellow, bottom row)
are concentrated in the west (Y1 haplotype), while others (blue, Arm1T) in the east.
http://1.bp.blogspot.com/-_mSQwcwmmik/UhzHzuryKTI/AAAAAAAAJE0/Po072BENEqg/s1600/ncomms3348-f1.jpg
{kind=link}
This is a nice demonstration of transfer of domesticated animals from Neolithic farmers to European hunter-gatherers. (Red=European, Yellow=Near Eastern).
The comings and goings of Near Eastern and European domestic pigs (Ottoni et al. 2012)
http://dienekes.blogspot.jp/2012/11/the-comings-and-goings-of-near-eastern.html
http://2.bp.blogspot.com/-yQYdErp2j2k/UK8tdYlo0OI/AAAAAAAAHmE/IchI7xSYLNQ/s1600/ottoni.png
{kind=link}
The left-right arrangement of the columns corresponds to a west-east longitude across West Asia.
It can be easily seen that some of the early domestic samples (yellow, bottom row)
are concentrated in the west (Y1 haplotype), while others (blue, Arm1T) in the east.
>>898
http://www.nature.com/ncomms/2013/130827/ncomms3348/fig_tab/ncomms3348_F1.html
Samples were recovered from Neolithic LBK, post-LBK and Mesolithic Ertebolle sites dated between 5500 and 4000 cal BC.
Each symbol corresponds to a single sample (triangle, square and circle).
Domestic (triangle) and wild (square) pigs discussed in the text are labelled; circles represent Sus specimens of unknown domestication status.
The red colour indicates the European haplotypes C and A, and yellow the Near Eastern haplotypes Y1 and Y2.
http://www.nature.com/ncomms/2013/130827/ncomms3348/fig_tab/ncomms3348_F1.html
Samples were recovered from Neolithic LBK, post-LBK and Mesolithic Ertebolle sites dated between 5500 and 4000 cal BC.
Each symbol corresponds to a single sample (triangle, square and circle).
Domestic (triangle) and wild (square) pigs discussed in the text are labelled; circles represent Sus specimens of unknown domestication status.
The red colour indicates the European haplotypes C and A, and yellow the Near Eastern haplotypes Y1 and Y2.
901 :世界@名無史さん:2014/02/03(月) 13:41:56.45 0
白人は顔が悪いヤツの方が多いぞ
白人が容姿端麗て言うヤツはハリウッド映画しか見てないんだろうな
白人が容姿端麗て言うヤツはハリウッド映画しか見てないんだろうな
19世紀に日本を訪ねたイギリス人
イザベラ・バード「日本奥地紀行」を読むと、
「日本人の容姿は醜い」という文章が何回も出てくる。
当時のイギリス人の美意識とは全く反していたのだろう>アジアン。
イザベラ・バード「日本奥地紀行」を読むと、
「日本人の容姿は醜い」という文章が何回も出てくる。
当時のイギリス人の美意識とは全く反していたのだろう>アジアン。
ルーニーとか凄い不細工だ
905 :世界@名無史さん:2014/02/03(月) 22:46:31.15 0
さて、900超えたらそろそろ終わるの?
突然落ちたりするんだっけ?
突然落ちたりするんだっけ?
906 :イタクァを作らなイカ?:2014/02/04(火) 21:39:48.35 0
この時誰もイカ娘の侵略の影に気づくものはイなカった
とりあえず、シリーズじゃない
世界の古代民族シリーズ
http://ci.nii.ac.jp/ncid/BA83939846
関連文献: 1件.書誌事項
世界の古代民族シリーズ
フェニキア人
グレン・E・マーコウ著 ; 片山陽子訳
創元社 2007.11 世界の古代民族シリーズ
所蔵館118館
世界の古代民族シリーズ
http://ci.nii.ac.jp/ncid/BA83939846
関連文献: 1件.書誌事項
世界の古代民族シリーズ
フェニキア人
グレン・E・マーコウ著 ; 片山陽子訳
創元社 2007.11 世界の古代民族シリーズ
所蔵館118館
カルタゴの母都市はテュロス
>>907 P68
カルタゴが政治・軍事勢力として浮上するのは、これまでのところ前6世紀の半ばとされる。
そのころからマゴ将軍とその子孫である後継者たち(マゴ家と呼ばれる)の指導で
征服と植民地拡大をねらった侵略的な軍事行動が開始されている。
サルディニア島やシチリア島のフェニキア人の植民地を守るために、
初めてそれらの島に軍事介入したのもだいたいそのころか、その少し前になる。
カルタゴの軍事力のまぎれもない証となるのが前535年の出来事である。
エトルリアの海軍と組んで、コルシカ島沖の海鮮でポカイアのギリシア人を破っている。
エトルリアはそのころティレニア同盟(トスカナ、ラティウム、カンパニアが参加していた)の先頭に立っていた国だったが、
カルタゴと商業的・政治的に密接に繋がりがあったというドラマティックな証拠品がある。
エトルリア語とフェニキア語で同じ内容が記された3枚の金製の奉納板が、
エトルリアの都市カエレの商港ピュルジで見つかっているのである。
カルタゴでエトルリアからの輸入品が多く見られることにも、二つの海上勢力の間の密接な通商関係が裏づけられるだろう。
実際この時代の地中海中部には、カルタゴの支配力がきわめて広範に及んでいたことが考古学調査で明らかにされている。
シチリア島のモティア、サルディニア島、それにカルタゴの最も古い海外植民地だったイビサ島などには、
文化の伝播と影響が色濃くみられることが発掘調査で裏づけられている。
>>907 P68
カルタゴが政治・軍事勢力として浮上するのは、これまでのところ前6世紀の半ばとされる。
そのころからマゴ将軍とその子孫である後継者たち(マゴ家と呼ばれる)の指導で
征服と植民地拡大をねらった侵略的な軍事行動が開始されている。
サルディニア島やシチリア島のフェニキア人の植民地を守るために、
初めてそれらの島に軍事介入したのもだいたいそのころか、その少し前になる。
カルタゴの軍事力のまぎれもない証となるのが前535年の出来事である。
エトルリアの海軍と組んで、コルシカ島沖の海鮮でポカイアのギリシア人を破っている。
エトルリアはそのころティレニア同盟(トスカナ、ラティウム、カンパニアが参加していた)の先頭に立っていた国だったが、
カルタゴと商業的・政治的に密接に繋がりがあったというドラマティックな証拠品がある。
エトルリア語とフェニキア語で同じ内容が記された3枚の金製の奉納板が、
エトルリアの都市カエレの商港ピュルジで見つかっているのである。
カルタゴでエトルリアからの輸入品が多く見られることにも、二つの海上勢力の間の密接な通商関係が裏づけられるだろう。
実際この時代の地中海中部には、カルタゴの支配力がきわめて広範に及んでいたことが考古学調査で明らかにされている。
シチリア島のモティア、サルディニア島、それにカルタゴの最も古い海外植民地だったイビサ島などには、
文化の伝播と影響が色濃くみられることが発掘調査で裏づけられている。
Q1. デーン人とは何か
Q2. エトルリア人とは何か
Q3. ピクト人とは何か
Q4. アーリア人とは何か
Q5. シュメール人とは何か
Q2. エトルリア人とは何か
Q3. ピクト人とは何か
Q4. アーリア人とは何か
Q5. シュメール人とは何か
なんかアレなのキタかも
http://dienekes.blogspot.jp/search?updated-min=2014-01-01T00:00:00%2B02:00&updated-max=2015-01-01T00:00:00%2B02:00&max-results=15
http://www.pnas.org/content/early/2014/01/29/1313787111.abstract
Ancient west Eurasian ancestry in southern and eastern Africa
Joseph K. Pickrell et al.
The history of southern Africa involved interactions between indigenous hunter?gatherers and a range of populations that moved into the region.
Here we use genome-wide genetic data to show that there are at least two admixture events
in the history of Khoisan populations (southern African hunter?gatherers and pastoralists who speak non-Bantu languages with click consonants).
One involved populations related to Niger?Congo-speaking African populations,
and the other introduced ancestry most closely related to west Eurasian (European or Middle Eastern) populations.
We date this latter admixture event to ?900?1,800 y ago and show that it had the largest demographic impact in Khoisan populations that speak Khoe?Kwadi languages.
A similar signal of west Eurasian ancestry is present throughout eastern Africa.
In particular, we also find evidence for two admixture events in the history of Kenyan, Tanzanian, and Ethiopian populations,
the earlier of which involved populations related to west Eurasians and which we date to ?2,700?3,300 y ago.
We reconstruct the allele frequencies of the putative west Eurasian population in eastern Africa and show that this population is a good proxy for the west Eurasian ancestry in southern Africa.
The most parsimonious explanation for these findings is that west Eurasian ancestry entered southern Africa indirectly through eastern Africa.
もしかして↓なのか
http://ja.wikipedia.org/wiki/%E3%83%8D%E3%82%B32%E4%B8%96
はたまた↓なのか
http://ja.wikipedia.org/wiki/%E3%83%8F%E3%83%88%E3%82%B7%E3%82%A7%E3%83%97%E3%82%B9%E3%83%88
http://dienekes.blogspot.jp/search?updated-min=2014-01-01T00:00:00%2B02:00&updated-max=2015-01-01T00:00:00%2B02:00&max-results=15
http://www.pnas.org/content/early/2014/01/29/1313787111.abstract
Ancient west Eurasian ancestry in southern and eastern Africa
Joseph K. Pickrell et al.
The history of southern Africa involved interactions between indigenous hunter?gatherers and a range of populations that moved into the region.
Here we use genome-wide genetic data to show that there are at least two admixture events
in the history of Khoisan populations (southern African hunter?gatherers and pastoralists who speak non-Bantu languages with click consonants).
One involved populations related to Niger?Congo-speaking African populations,
and the other introduced ancestry most closely related to west Eurasian (European or Middle Eastern) populations.
We date this latter admixture event to ?900?1,800 y ago and show that it had the largest demographic impact in Khoisan populations that speak Khoe?Kwadi languages.
A similar signal of west Eurasian ancestry is present throughout eastern Africa.
In particular, we also find evidence for two admixture events in the history of Kenyan, Tanzanian, and Ethiopian populations,
the earlier of which involved populations related to west Eurasians and which we date to ?2,700?3,300 y ago.
We reconstruct the allele frequencies of the putative west Eurasian population in eastern Africa and show that this population is a good proxy for the west Eurasian ancestry in southern Africa.
The most parsimonious explanation for these findings is that west Eurasian ancestry entered southern Africa indirectly through eastern Africa.
もしかして↓なのか
http://ja.wikipedia.org/wiki/%E3%83%8D%E3%82%B32%E4%B8%96
はたまた↓なのか
http://ja.wikipedia.org/wiki/%E3%83%8F%E3%83%88%E3%82%B7%E3%82%A7%E3%83%97%E3%82%B9%E3%83%88
911 :世界@名無史さん:2014/02/05(水) 22:40:46.14 0
マサイ族のY-DNAは50%がE1b1bで、Mt-DNAでも12.5%がMだけど、さらに南下してコイサンにまで中東?ヨーロッパ?の影響は大航海時代より前に及んでたのか。
コイコイ、いわゆるホッテントットに牧畜を教えて家畜を与えたのがこの時の混交かな?900〜1800年前のどこかと言うと相当幅があるけど。更にそれ以前にニジェール・コンゴ語族と混血か
これらの混血と牧畜を身に着けてからアフリカ南部に行ったのかな?狩猟採集民のままだったサン(ブッシュマン)にはこの混血の影響は及んだのかどうか。
コイコイ、いわゆるホッテントットに牧畜を教えて家畜を与えたのがこの時の混交かな?900〜1800年前のどこかと言うと相当幅があるけど。更にそれ以前にニジェール・コンゴ語族と混血か
これらの混血と牧畜を身に着けてからアフリカ南部に行ったのかな?狩猟採集民のままだったサン(ブッシュマン)にはこの混血の影響は及んだのかどうか。
>>911
→E. Africa
Afar
Amhara
Anuanak
AriBlacksmith
AriCultivator
ESomali
Gumuz
Hadza
Luhya
Maasai
Oromo
Sandawe
Somali
Tygray
Wolayta
→S. Africa
!Xuun
GuiGana
Juhoansi
Khomani
Khwe
Nama
だけど詳しくわからんww
Westユーラシアは、むしろ地中海人で、
サルディニア、キプロス、トスカーナあたりが想定されてそう
→E. Africa
Afar
Amhara
Anuanak
AriBlacksmith
AriCultivator
ESomali
Gumuz
Hadza
Luhya
Maasai
Oromo
Sandawe
Somali
Tygray
Wolayta
→S. Africa
!Xuun
GuiGana
Juhoansi
Khomani
Khwe
Nama
だけど詳しくわからんww
Westユーラシアは、むしろ地中海人で、
サルディニア、キプロス、トスカーナあたりが想定されてそう
>>91>>912
これ以外で重要そうなのは、イエメンのユダヤ教徒なんだけど
全般にエチオピア周辺に強いマーカーが
さて、プントは何処にあったんだろうか?
http://ja.wikipedia.org/wiki/%E3%83%97%E3%83%B3%E3%83%88%E5%9B%BD
これ以外で重要そうなのは、イエメンのユダヤ教徒なんだけど
全般にエチオピア周辺に強いマーカーが
さて、プントは何処にあったんだろうか?
http://ja.wikipedia.org/wiki/%E3%83%97%E3%83%B3%E3%83%88%E5%9B%BD
914 :世界@名無史さん:2014/02/06(木) 03:06:21.24 0
プント=バントゥー説は否定されているようだが、それではプントとはなんだったのか
http://upload.wikimedia.org/wikipedia/commons/7/75/Queen_of_punt.jpg プント女王の肖像画
筋ジストロフィーや象皮病とも言われるが、これを連想した
realhistoryww.com/world_history/ancient/Misc/Prehistoric_Art/Steatopygia_Female.jpg
コイコイ女性に多く見られると言うが
アフリカの角に謎のコイサン国家が存在したのか?文明レベルでないにしても定住社会が存在したのか
http://upload.wikimedia.org/wikipedia/commons/7/75/Queen_of_punt.jpg プント女王の肖像画
{kind=link}
筋ジストロフィーや象皮病とも言われるが、これを連想した
realhistoryww.com/world_history/ancient/Misc/Prehistoric_Art/Steatopygia_Female.jpg
コイコイ女性に多く見られると言うが
アフリカの角に謎のコイサン国家が存在したのか?文明レベルでないにしても定住社会が存在したのか
白人の頭身、手足のバランスは黄金比によってなりたってるに違いないため美しいのであろう
>>604>>698
>>876>>911
>>910
アフリカにおける農耕民と採集狩猟民との関係
http://sicambre.at.webry.info/201402/article_6.html
アフリカ中西部では5000年前頃に農業が始まり、サハラ砂漠以南のアフリカ全体で農耕型生活様式が拡大していきました。
しかし、採集狩猟民として生活し続けた集団も存在します。
この研究によると、農耕民と採集狩猟民との間の遺伝子流動は過去1000年間に限られていたことが判明したそうです。
以下は『ネイチャー』の日本語サイトからの引用です。
【進化】熱帯雨林の狩猟採集民に対する農業の影響
アフリカにおいて農耕民と農耕民が持ち込んだ農業が熱帯雨林の狩猟採集民に与えた影響を調べた研究の結果を報告する論文が掲載される。
今回の研究は、農業が熱帯雨林の狩猟採集民と農耕民の集団に与えた影響を明らかにし、この2つの集団の間で最近起こった遺伝的交換を明確に示している。
アフリカ中西部では、今からおよそ5000年前に農業が出現し、それによって、サハラ以南のアフリカ全体で、運動量の少ない農耕型生活様式の拡大が始まった。
しかし、一部の熱帯雨林の狩猟採集民集団は、移動集団の生活を続けた。
農業文化と技術の拡散が熱帯雨林の狩猟採集民の人口統計学的歴史に及ぼした影響の全容は、ほとんど解明されないままになっている。
今回、Lluis Quintana-Murciたちは、サハラ以南のアフリカにおいて、現代の熱帯雨林の狩猟採集民と農耕民のゲノム規模の一塩基多型データを生成し、
この2つの集団について、遺伝的多様性と集団間の進化的関係を調べた。
その結果、この2集団間の遺伝子流動が、過去1000年間に限られていたことが判明した。
このことは、2集団間での当初の相互作用が、社会経済的発想(例えば、道具や植物栽培技術)の交換に限定されていたことを示唆している。
さらに、今回明らかになったゲノムの特徴は、アフリカでの農耕民集団の増加が農業の出現の前から起こっており、
従って、農耕民の増加は農業の出現によるものとはいえないことを示唆している。
参考文献:
http://dx.doi.org/10.1038/ncomms4163
>>876>>911
>>910
アフリカにおける農耕民と採集狩猟民との関係
http://sicambre.at.webry.info/201402/article_6.html
アフリカ中西部では5000年前頃に農業が始まり、サハラ砂漠以南のアフリカ全体で農耕型生活様式が拡大していきました。
しかし、採集狩猟民として生活し続けた集団も存在します。
この研究によると、農耕民と採集狩猟民との間の遺伝子流動は過去1000年間に限られていたことが判明したそうです。
以下は『ネイチャー』の日本語サイトからの引用です。
【進化】熱帯雨林の狩猟採集民に対する農業の影響
アフリカにおいて農耕民と農耕民が持ち込んだ農業が熱帯雨林の狩猟採集民に与えた影響を調べた研究の結果を報告する論文が掲載される。
今回の研究は、農業が熱帯雨林の狩猟採集民と農耕民の集団に与えた影響を明らかにし、この2つの集団の間で最近起こった遺伝的交換を明確に示している。
アフリカ中西部では、今からおよそ5000年前に農業が出現し、それによって、サハラ以南のアフリカ全体で、運動量の少ない農耕型生活様式の拡大が始まった。
しかし、一部の熱帯雨林の狩猟採集民集団は、移動集団の生活を続けた。
農業文化と技術の拡散が熱帯雨林の狩猟採集民の人口統計学的歴史に及ぼした影響の全容は、ほとんど解明されないままになっている。
今回、Lluis Quintana-Murciたちは、サハラ以南のアフリカにおいて、現代の熱帯雨林の狩猟採集民と農耕民のゲノム規模の一塩基多型データを生成し、
この2つの集団について、遺伝的多様性と集団間の進化的関係を調べた。
その結果、この2集団間の遺伝子流動が、過去1000年間に限られていたことが判明した。
このことは、2集団間での当初の相互作用が、社会経済的発想(例えば、道具や植物栽培技術)の交換に限定されていたことを示唆している。
さらに、今回明らかになったゲノムの特徴は、アフリカでの農耕民集団の増加が農業の出現の前から起こっており、
従って、農耕民の増加は農業の出現によるものとはいえないことを示唆している。
参考文献:
http://dx.doi.org/10.1038/ncomms4163
>>917
http://www.nature.com/ncomms/2014/140204/ncomms4163/fig_tab/ncomms4163_ft.html
これは完全にアフリカ話なのでやりすぎちゃったかもね
Figure 1: Genome-wide structure of RHG and AGR populations.
http://www.nature.com/ncomms/2014/140204/ncomms4163/fig_tab/ncomms4163_F1.html
ここに地図が載ってるけど
http://www.nature.com/ncomms/2014/140204/ncomms4163/fig_tab/ncomms4163_ft.html
これは完全にアフリカ話なのでやりすぎちゃったかもね
Figure 1: Genome-wide structure of RHG and AGR populations.
http://www.nature.com/ncomms/2014/140204/ncomms4163/fig_tab/ncomms4163_F1.html
ここに地図が載ってるけど
919 :世界@名無史さん:2014/02/06(木) 23:54:22.19 0
ピグミーと他の集団との遺伝的関係か。少なくとも10世紀ころまではピグミー独自の言語が保たれてたってことか
>アフリカでの農耕民集団の増加が農業の出現の前から起こっており、従って、農耕民の増加は農業の出現によるものとはいえないことを示唆している。
農業を始めたから人口が増えたんじゃなく、人口が増えたから農業を始めざるを得なくなったってことか?しばらくは増える人口を狩猟採集でも養えたけど。つまりは世界の農耕の起源地の1つってこと?
アフリカイネの栽培化はたしかにそうだな。でもその最初の狩猟民時代の人口増加のきっかけってなんだろう
なんかどんどんヨーロッパから離れてってるw
>アフリカでの農耕民集団の増加が農業の出現の前から起こっており、従って、農耕民の増加は農業の出現によるものとはいえないことを示唆している。
農業を始めたから人口が増えたんじゃなく、人口が増えたから農業を始めざるを得なくなったってことか?しばらくは増える人口を狩猟採集でも養えたけど。つまりは世界の農耕の起源地の1つってこと?
アフリカイネの栽培化はたしかにそうだな。でもその最初の狩猟民時代の人口増加のきっかけってなんだろう
なんかどんどんヨーロッパから離れてってるw
>>863>>892>>909
Genetic Diversity among Ancient Nordic Populations
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0011898
Using established criteria for work with fossil DNA we have analysed mitochondrial DNA from 92 individuals from 18 locations in Denmark ranging in time from the Mesolithic to the Medieval Age.
Unequivocal assignment of mtDNA haplotypes was possible for 56 of the ancient individuals;
however, the success rate varied substantially between sites; the highest rates were obtained with untouched, freshly excavated material,
whereas heavy handling, archeological preservation and storage for many years influenced the ability to obtain authentic endogenic DNA.
While the nucleotide diversity at two locations was similar to that among extant Danes, the diversity at four sites was considerably higher.
This supports previous observations for ancient Britons. The overall occurrence of haplogroups did not deviate from extant Scandinavians,
however, haplogroup I was significantly more frequent among the ancient Danes (average 13%) than among extant Danes and Scandinavians (~2.5%) as well as among other ancient population samples reported.
Haplogroup I could therefore have been an ancient Southern Scandinavian type “diluted” by later immigration events.
Interestingly, the two Neolithic samples (4,200 YBP, Bell Beaker culture) that were typed were haplogroup U4 and U5a, respectively, and the single Bronze Age sample (3,300?3,500 YBP) was haplogroup U4.
These two haplogroups have been associated with the Mesolithic populations of Central and Northern Europe.
Therefore, at least for Southern Scandinavia, our findings do not support a possible replacement of a haplogroup U dominated hunter-gatherer population by a more haplogroup diverse Neolithic Culture.
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0011898
Genetic Diversity among Ancient Nordic Populations
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0011898
Using established criteria for work with fossil DNA we have analysed mitochondrial DNA from 92 individuals from 18 locations in Denmark ranging in time from the Mesolithic to the Medieval Age.
Unequivocal assignment of mtDNA haplotypes was possible for 56 of the ancient individuals;
however, the success rate varied substantially between sites; the highest rates were obtained with untouched, freshly excavated material,
whereas heavy handling, archeological preservation and storage for many years influenced the ability to obtain authentic endogenic DNA.
While the nucleotide diversity at two locations was similar to that among extant Danes, the diversity at four sites was considerably higher.
This supports previous observations for ancient Britons. The overall occurrence of haplogroups did not deviate from extant Scandinavians,
however, haplogroup I was significantly more frequent among the ancient Danes (average 13%) than among extant Danes and Scandinavians (~2.5%) as well as among other ancient population samples reported.
Haplogroup I could therefore have been an ancient Southern Scandinavian type “diluted” by later immigration events.
Interestingly, the two Neolithic samples (4,200 YBP, Bell Beaker culture) that were typed were haplogroup U4 and U5a, respectively, and the single Bronze Age sample (3,300?3,500 YBP) was haplogroup U4.
These two haplogroups have been associated with the Mesolithic populations of Central and Northern Europe.
Therefore, at least for Southern Scandinavia, our findings do not support a possible replacement of a haplogroup U dominated hunter-gatherer population by a more haplogroup diverse Neolithic Culture.
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0011898
921 :世界@名無史さん:2014/02/07(金) 00:03:54.78 0
>>910 アフリカ中部に見られるこの傾向は、アフリカ大陸の南部で起きたことと全く異なっていると論文は述べている。
南部では、影響力の強い農耕民族と狩猟採集民族サン(San)との遭遇が「遺伝子の速やかな交換をもたらした」という。
-少数民族ピグミーの歴史、遺伝子で解明 仏研究 AFPBPニュース 2014/2/5 より
サンにもコイコイ同様外部由来の遺伝子が受け継がれたか。それが比較的後世のバントゥ系の南部アフリカ進入時か、まだサンの祖先が東アフリカにいた頃かはわからないけど。
でも長く分離を保ったのに結局は言語が失われたピグミーと、速やかに混血しても言語を保ったコイサンとは、皮肉な展開だな
南部では、影響力の強い農耕民族と狩猟採集民族サン(San)との遭遇が「遺伝子の速やかな交換をもたらした」という。
-少数民族ピグミーの歴史、遺伝子で解明 仏研究 AFPBPニュース 2014/2/5 より
サンにもコイコイ同様外部由来の遺伝子が受け継がれたか。それが比較的後世のバントゥ系の南部アフリカ進入時か、まだサンの祖先が東アフリカにいた頃かはわからないけど。
でも長く分離を保ったのに結局は言語が失われたピグミーと、速やかに混血しても言語を保ったコイサンとは、皮肉な展開だな
>>919>>921
アフリカはよくわからんww
気候変動が農耕をもたらした説は古くからあるけど
>>877
サハラ砂漠で古代の墓地を発見 よみがえる緑豊かな時代
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=6667009
サハラ砂漠で石器時代の最大最古の墓地を発見したのは現代の恐竜探検家だった。
ナショナル ジオグラフィック協会付き探検家であるシカゴ大学の古生物学者、ポール・セレノ氏の調査チームは、2000年にニジェール北部の荒々しい砂丘で恐竜の骨を探していた。
かつては湖の岸辺だったその場所で岩を調べていたところ、思いがけず墓地に遭遇したのだった。
最終的に調査チームは、5000年の期間にまたがり大きく異なる2つの文明に属する200個の墓を発掘した。このような墓地が発見されるのは初めてのことだ。
セレノ氏が最近発表した研究結果によると、ゴベロと呼ばれるこの地域は、
キフィアン文明(紀元前7700〜6200年)とテネリアン文明(紀元前5200〜2500年)の居住地と墓が保存された貴重な場所だという。
緑豊かなサハラが乾燥して世界最大の砂漠となった完新世時代に、人々がどのように生活し、死者をどのように埋葬していたのかを解明する上で新たな扉が開かれた。
セレノ氏のチームによると、約1万2000年前に地球の軌道の傾きやそのほかの環境的要因が重なって、サハラに強烈なモンスーンが吹き荒れた。
その結果、砂漠が緑化し、居住地を求めて人間が移動してきたという。
湿潤な時期に狩猟採集民のキフィアン人がこの地域を占有していたことは既に分かっていた。
紀元前6200〜5200年頃には、その時代で最も激しい気候変動が起きて土地が乾燥し、居住民は移動を余儀なくされた。
間もなくして2番目のグループのテネリアン人がこの地に到着した。
アフリカはよくわからんww
気候変動が農耕をもたらした説は古くからあるけど
>>877
サハラ砂漠で古代の墓地を発見 よみがえる緑豊かな時代
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=6667009
サハラ砂漠で石器時代の最大最古の墓地を発見したのは現代の恐竜探検家だった。
ナショナル ジオグラフィック協会付き探検家であるシカゴ大学の古生物学者、ポール・セレノ氏の調査チームは、2000年にニジェール北部の荒々しい砂丘で恐竜の骨を探していた。
かつては湖の岸辺だったその場所で岩を調べていたところ、思いがけず墓地に遭遇したのだった。
最終的に調査チームは、5000年の期間にまたがり大きく異なる2つの文明に属する200個の墓を発掘した。このような墓地が発見されるのは初めてのことだ。
セレノ氏が最近発表した研究結果によると、ゴベロと呼ばれるこの地域は、
キフィアン文明(紀元前7700〜6200年)とテネリアン文明(紀元前5200〜2500年)の居住地と墓が保存された貴重な場所だという。
緑豊かなサハラが乾燥して世界最大の砂漠となった完新世時代に、人々がどのように生活し、死者をどのように埋葬していたのかを解明する上で新たな扉が開かれた。
セレノ氏のチームによると、約1万2000年前に地球の軌道の傾きやそのほかの環境的要因が重なって、サハラに強烈なモンスーンが吹き荒れた。
その結果、砂漠が緑化し、居住地を求めて人間が移動してきたという。
湿潤な時期に狩猟採集民のキフィアン人がこの地域を占有していたことは既に分かっていた。
紀元前6200〜5200年頃には、その時代で最も激しい気候変動が起きて土地が乾燥し、居住民は移動を余儀なくされた。
間もなくして2番目のグループのテネリアン人がこの地に到着した。
>>922
以前はこうした人口移動の証拠としては道具などの人工物しか存在せず、分析の対象となる人骨が見つかっていなかった。
しかし、その状況を今回の発見が変えてくれた。出土した骨を放射性炭素年代測定で調べた結果、古代サハラの「顕著な記録」がもたらされたとセレノ氏は話す。
「人々がかつて暮らしていた緑豊かなサハラの記録が砂丘に刻まれていたのだ」。
出土品の中には、現在もアフリカ南部に生息するイボイノシシやクロコダイル、カバなどの野生動物の骨や牙、
陶器、象牙や貝殻の装飾品といった副葬品もあり、葬儀の様子を解明する手掛かりとなっている。
おそらく最も印象的なのは、2006年に発見された「石器時代の抱擁」と呼ばれる遺骨だろう。テネリアン人の女性と2人の子どもが向き合って腕でポーズを取り、手を握り合っている。
骨格の下にある花粉の残留物は、死者が花のベッドの上に横たわっていたことを示している。
セレノ氏は「ここは画期的な墓地だ。先史時代のこのような場所はほかには見つかっていない」と話している。
この遺跡から、ほとんど研究が進んでいない文明についてさまざまな洞察を進めることができるが、それでも残る謎があるとセレノ氏は言う。
最大の謎は、テネリアン人はどうやってキフィアン人の遺体を傷つけずに隣りに新しい墓を掘ることができたのかということだ。これは「驚くべき離れ技」だという。
しかし、なぜこの2つの文明がガベロで死者の埋葬をしたのかは明らかである。
それは、現代で言えばデイトナビーチのように、完新世には魚と動物が集まる理想的な場所である湖畔に位置した最も望ましい土地だったと、セレノ氏は語っている。
以前はこうした人口移動の証拠としては道具などの人工物しか存在せず、分析の対象となる人骨が見つかっていなかった。
しかし、その状況を今回の発見が変えてくれた。出土した骨を放射性炭素年代測定で調べた結果、古代サハラの「顕著な記録」がもたらされたとセレノ氏は話す。
「人々がかつて暮らしていた緑豊かなサハラの記録が砂丘に刻まれていたのだ」。
出土品の中には、現在もアフリカ南部に生息するイボイノシシやクロコダイル、カバなどの野生動物の骨や牙、
陶器、象牙や貝殻の装飾品といった副葬品もあり、葬儀の様子を解明する手掛かりとなっている。
おそらく最も印象的なのは、2006年に発見された「石器時代の抱擁」と呼ばれる遺骨だろう。テネリアン人の女性と2人の子どもが向き合って腕でポーズを取り、手を握り合っている。
骨格の下にある花粉の残留物は、死者が花のベッドの上に横たわっていたことを示している。
セレノ氏は「ここは画期的な墓地だ。先史時代のこのような場所はほかには見つかっていない」と話している。
この遺跡から、ほとんど研究が進んでいない文明についてさまざまな洞察を進めることができるが、それでも残る謎があるとセレノ氏は言う。
最大の謎は、テネリアン人はどうやってキフィアン人の遺体を傷つけずに隣りに新しい墓を掘ることができたのかということだ。これは「驚くべき離れ技」だという。
しかし、なぜこの2つの文明がガベロで死者の埋葬をしたのかは明らかである。
それは、現代で言えばデイトナビーチのように、完新世には魚と動物が集まる理想的な場所である湖畔に位置した最も望ましい土地だったと、セレノ氏は語っている。
トカラ語
http://ja.wikipedia.org/wiki/%E3%83%88%E3%82%AB%E3%83%A9%E8%AA%9E
トカラ語(トカラご、トハラ語、英:Tocharian languages)は、
現在の中華人民共和国新疆ウイグル自治区(東トルキスタン)のタリム盆地北縁(天山南路)にあたる地域で8世紀頃まで話され、
ブラーフミー文字に似たトカラ文字(英語版)で書いた写本が残っている言語。
インド・ヨーロッパ語族に属し、独立した語派「トカラ語派」を形成する。膠着語的な性格を有していたことが分かっている。
概説[編集]
ブラーフミー文字で書かれた西暦500〜700年頃の文献が見つかっている。
8世紀頃までに古テュルク語(英語版)の話者集団に吸収され、死滅したと考えられている。
なお、かつて紀元前後にストラボンが記した「トカロイ」、
中国史書が伝える「吐呼羅国」[1]、「吐火羅国」[2]、7世紀に玄奘が記した「覩貨邏国」[3]といった、
いわゆる「トハラ人」および「トハリスタン」とは区別する[4]立場もある。
音声面ではインド・ヨーロッパ祖語の *? 音を k として保持している、いわゆるケントゥム語群の特性を有し、
同語群の他の言語から地理的に大きく離れていることから特異視されることもある。
起源[編集]
楼蘭などでは紀元後にカロシュティー文字で書かれるガンダーリ語(英語版)(インド語派。ヴェーダ語や近縁の他のプラークリットから派生したと考えられている)が用いられていたが、
その中にもトカラ語に類似した単語が散見されるため、古くはタリム盆地の多くの地域でトカラ語派の言語を話していたと思われる。
トカラ語派を話す人たちはミイラのDNA調査の結果、後述のアファナシェヴォ文化を担った集団と非常に近く、
個体の7割が南シベリアに特徴的なミトコンドリアDNAを持っていたため、北方から南下して来たことが明らかになった。
タリム盆地では紀元前2千年前後といわれる遺跡(「楼蘭の美女」などのミイラで有名)が発見されており、
これらは上述の検査や葬制の類似から、紀元前3千年紀の青銅器時代初期に北方のアルタイ山脈方面に栄えたアファナシェヴォ文化人が
西からインド系集団に征服されながら南下してタリム盆地へ移り住んだことが判明しており、アファナシェヴォ文化はトカラ語と強い関連があると考えられている。
http://ja.wikipedia.org/wiki/%E3%83%88%E3%82%AB%E3%83%A9%E8%AA%9E
トカラ語(トカラご、トハラ語、英:Tocharian languages)は、
現在の中華人民共和国新疆ウイグル自治区(東トルキスタン)のタリム盆地北縁(天山南路)にあたる地域で8世紀頃まで話され、
ブラーフミー文字に似たトカラ文字(英語版)で書いた写本が残っている言語。
インド・ヨーロッパ語族に属し、独立した語派「トカラ語派」を形成する。膠着語的な性格を有していたことが分かっている。
概説[編集]
ブラーフミー文字で書かれた西暦500〜700年頃の文献が見つかっている。
8世紀頃までに古テュルク語(英語版)の話者集団に吸収され、死滅したと考えられている。
なお、かつて紀元前後にストラボンが記した「トカロイ」、
中国史書が伝える「吐呼羅国」[1]、「吐火羅国」[2]、7世紀に玄奘が記した「覩貨邏国」[3]といった、
いわゆる「トハラ人」および「トハリスタン」とは区別する[4]立場もある。
音声面ではインド・ヨーロッパ祖語の *? 音を k として保持している、いわゆるケントゥム語群の特性を有し、
同語群の他の言語から地理的に大きく離れていることから特異視されることもある。
起源[編集]
楼蘭などでは紀元後にカロシュティー文字で書かれるガンダーリ語(英語版)(インド語派。ヴェーダ語や近縁の他のプラークリットから派生したと考えられている)が用いられていたが、
その中にもトカラ語に類似した単語が散見されるため、古くはタリム盆地の多くの地域でトカラ語派の言語を話していたと思われる。
トカラ語派を話す人たちはミイラのDNA調査の結果、後述のアファナシェヴォ文化を担った集団と非常に近く、
個体の7割が南シベリアに特徴的なミトコンドリアDNAを持っていたため、北方から南下して来たことが明らかになった。
タリム盆地では紀元前2千年前後といわれる遺跡(「楼蘭の美女」などのミイラで有名)が発見されており、
これらは上述の検査や葬制の類似から、紀元前3千年紀の青銅器時代初期に北方のアルタイ山脈方面に栄えたアファナシェヴォ文化人が
西からインド系集団に征服されながら南下してタリム盆地へ移り住んだことが判明しており、アファナシェヴォ文化はトカラ語と強い関連があると考えられている。
アファナシェヴォ文化
アファナシェヴォ文化(-ぶんか、Afanasevo culture)は、紀元前3500年から2500年頃、中央アジア北東部からシベリア南部にかけて栄えた文化で、
銅器時代後期ないし青銅器時代前期に当たる。シベリア・ミヌシンスク盆地のアファナシェヴォで最初に発掘調査されたが、
現在のモンゴル西部、新疆ウイグル自治区北部、カザフスタン中東部にまで広がっていた。
さらにタジキスタン・アラル海方面の文化も関連があるとされる。
生活様式は半遊牧的牧畜と考えられる。牛、羊または山羊、馬を飼育したが、野生動物の狩猟も行ったようであり、ともに出土する例が多い。
葬制に特徴があり、墓槨は円錐形または矩形、多くは仰臥位に埋葬されて赤土がかけられ、環状列石が立てられた。集落遺跡も多数発見され、金属製品と車が発見されている。
人種的にはコーカソイドであり、葬制には西方ポンティック・ステップ地帯のヤムナ文化(竪穴墓文化)などとの類似点が多い。
ギンブタスによって提唱されたクルガン仮説(ヤムナ文化をクルガンIV期とする)によれば、これらは原印欧民族あるいはその後継者によるものとされる。
アファナシェヴォ文化が印欧語族によるものとすれば、その中で非常に古い時代に最も北東に位置した民族ということになる。
以上に関連して、トカラ語派(紀元後まで南方のタリム盆地などで話され文字に書かれた)の源流であるとする説も有力である。
これについては、紀元前1800年頃のタリム盆地における葬制(「楼蘭の美女」などのミイラで有名)や、
家畜と野生動物の骨がともに出土するなどの共通点が指摘されているが、直接的には証明されていない。
アファナシェヴォ文化の後、南シベリアではモンゴロイドを主体とするオクネフ文化が続く。
さらに西方の南ウラル方面に現れたアンドロノヴォ文化(インド・イラン語派に関係があるとする説が有力)がこの地域にまで広がる。
アファナシェヴォ文化(-ぶんか、Afanasevo culture)は、紀元前3500年から2500年頃、中央アジア北東部からシベリア南部にかけて栄えた文化で、
銅器時代後期ないし青銅器時代前期に当たる。シベリア・ミヌシンスク盆地のアファナシェヴォで最初に発掘調査されたが、
現在のモンゴル西部、新疆ウイグル自治区北部、カザフスタン中東部にまで広がっていた。
さらにタジキスタン・アラル海方面の文化も関連があるとされる。
生活様式は半遊牧的牧畜と考えられる。牛、羊または山羊、馬を飼育したが、野生動物の狩猟も行ったようであり、ともに出土する例が多い。
葬制に特徴があり、墓槨は円錐形または矩形、多くは仰臥位に埋葬されて赤土がかけられ、環状列石が立てられた。集落遺跡も多数発見され、金属製品と車が発見されている。
人種的にはコーカソイドであり、葬制には西方ポンティック・ステップ地帯のヤムナ文化(竪穴墓文化)などとの類似点が多い。
ギンブタスによって提唱されたクルガン仮説(ヤムナ文化をクルガンIV期とする)によれば、これらは原印欧民族あるいはその後継者によるものとされる。
アファナシェヴォ文化が印欧語族によるものとすれば、その中で非常に古い時代に最も北東に位置した民族ということになる。
以上に関連して、トカラ語派(紀元後まで南方のタリム盆地などで話され文字に書かれた)の源流であるとする説も有力である。
これについては、紀元前1800年頃のタリム盆地における葬制(「楼蘭の美女」などのミイラで有名)や、
家畜と野生動物の骨がともに出土するなどの共通点が指摘されているが、直接的には証明されていない。
アファナシェヴォ文化の後、南シベリアではモンゴロイドを主体とするオクネフ文化が続く。
さらに西方の南ウラル方面に現れたアンドロノヴォ文化(インド・イラン語派に関係があるとする説が有力)がこの地域にまで広がる。
ケントゥム語派
http://ja.wikipedia.org/wiki/%E3%82%B1%E3%83%B3%E3%83%88%E3%82%A5%E3%83%A0%E8%AA%9E%E6%B4%BE
ケントゥム語派に属するのはイタリック語派、ケルト語派、ゲルマン語派、ギリシャ語、アルバニア語のみで、
インド・ヨーロッパ語族の分布の中で西側に位置しているとされ、ケントゥム語派とサテム語派の間には断絶があるとされてきた。
ところが20世紀に入って新たに発見された死語であるアナトリア語派とトカラ語派は、インド・ヨーロッパ語族の分布の中で東に位置していた。
とくにトカラ語派は間にサテム語派の分布域を挟んではるかかなたの東方に位置している。
外部リンクによれば、印欧語族はもとはケントゥム語派の発音を用いており
後の時代にその分布の中央部(おそらくコーカサス地方)で現在のサテム語派が用いる発音の流行が発生し、
その新しい音声の流行が周囲の各言語集団へと次々に伝播していった。
この発音上の流行を「サテム化(satemization)」という。
このためケントゥム語派の用いる古い発音が印欧語族の分布の東と西にそれぞれまとまって残ったとされる。
とくにスラヴ語派とバルト語派はどちらもサテム語派に属するものの、
同時にケントゥム語派の音声的特徴も残しており(スラヴ語派はバルト語派よりもケントゥム語群の名残が強い)、
またこのスラヴ語派とバルト語派は文法的にはゲルマン語派(ケントゥム語派に属する)との間で明確な共通性があるため、
スラヴ語派、バルト語派、ゲルマン語派の3つの言語の共通祖語(インド・ヨーロッパ祖語の北西語群)を想定する学説も有力となってきている[1][2]。
http://ja.wikipedia.org/wiki/%E3%82%B1%E3%83%B3%E3%83%88%E3%82%A5%E3%83%A0%E8%AA%9E%E6%B4%BE
ケントゥム語派に属するのはイタリック語派、ケルト語派、ゲルマン語派、ギリシャ語、アルバニア語のみで、
インド・ヨーロッパ語族の分布の中で西側に位置しているとされ、ケントゥム語派とサテム語派の間には断絶があるとされてきた。
ところが20世紀に入って新たに発見された死語であるアナトリア語派とトカラ語派は、インド・ヨーロッパ語族の分布の中で東に位置していた。
とくにトカラ語派は間にサテム語派の分布域を挟んではるかかなたの東方に位置している。
外部リンクによれば、印欧語族はもとはケントゥム語派の発音を用いており
後の時代にその分布の中央部(おそらくコーカサス地方)で現在のサテム語派が用いる発音の流行が発生し、
その新しい音声の流行が周囲の各言語集団へと次々に伝播していった。
この発音上の流行を「サテム化(satemization)」という。
このためケントゥム語派の用いる古い発音が印欧語族の分布の東と西にそれぞれまとまって残ったとされる。
とくにスラヴ語派とバルト語派はどちらもサテム語派に属するものの、
同時にケントゥム語派の音声的特徴も残しており(スラヴ語派はバルト語派よりもケントゥム語群の名残が強い)、
またこのスラヴ語派とバルト語派は文法的にはゲルマン語派(ケントゥム語派に属する)との間で明確な共通性があるため、
スラヴ語派、バルト語派、ゲルマン語派の3つの言語の共通祖語(インド・ヨーロッパ祖語の北西語群)を想定する学説も有力となってきている[1][2]。
インド・ヨーロッパ古文献に関する覚え書き
http://www.hmn.bun.kyoto-u.ac.jp/report/2-pdf/4_bungaku1/4_02.pdf
この点に注目するなら、従来の系統樹モデルとは異なる言語の分岐モデルが考えられる。
それは共通祖語からアナトリア語派がまず離脱し、それに続いてトカラ語、
さらにイタリック語派、ケルト語派が離脱したが、中央ヨーロッパに残った諸言語はなお言語共同体としての
共通の特徴を持っていたという見方である。
そしてさらに後の時期に、残った諸言語のうち東側グループ
(伝統的にサタム語群)とよばれているグループ、
すなわちインド語、イラン語、スラブ語、バルト語、アルメニア語、アルバニア語)に*kからの歯擦音への変化が起こった。
この見方をとれば*kからの歯擦音への変化は早い時期に分岐した
アナトリア語派、トカラ語、イタリック語派、ケルト語派、
それに残りの中央ヨーロッパにとどまった諸言語のうち
西側のグループ(ギリシア語とゲルマン諸語)には及ばなかったことになる。
http://www.hmn.bun.kyoto-u.ac.jp/report/2-pdf/4_bungaku1/4_02.pdf
この点に注目するなら、従来の系統樹モデルとは異なる言語の分岐モデルが考えられる。
それは共通祖語からアナトリア語派がまず離脱し、それに続いてトカラ語、
さらにイタリック語派、ケルト語派が離脱したが、中央ヨーロッパに残った諸言語はなお言語共同体としての
共通の特徴を持っていたという見方である。
そしてさらに後の時期に、残った諸言語のうち東側グループ
(伝統的にサタム語群)とよばれているグループ、
すなわちインド語、イラン語、スラブ語、バルト語、アルメニア語、アルバニア語)に*kからの歯擦音への変化が起こった。
この見方をとれば*kからの歯擦音への変化は早い時期に分岐した
アナトリア語派、トカラ語、イタリック語派、ケルト語派、
それに残りの中央ヨーロッパにとどまった諸言語のうち
西側のグループ(ギリシア語とゲルマン諸語)には及ばなかったことになる。
アナトリア語派
http://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%8A%E3%83%88%E3%83%AA%E3%82%A2%E8%AA%9E%E6%B4%BE
アナトリア語派(アナトリアごは、Anatolian languages)とは古代小アジア(アナトリア:現在のトルコ)で話されていたインド・ヨーロッパ語族の言語群で、
すべて死語である。中でもヒッタイト語の資料が多く最もよく研究されている。
その周辺で使われた言語もいくつか知られるが、これらについては資料が少ない。
起源
アナトリア語派はインド・ヨーロッパ祖語(印欧祖語)から最も早期に分かれたとの説が有力で、
一部では「インド・ヒッタイト語族」という言い方もされる。
この時期については紀元前4千年紀半ばとするクルガン仮説があるが、さらに古いとする説(レンフリュー説、また語彙統計学による報告[1])もある。
クルガン仮説によれば、初期のアナトリア語派話者が黒海の北からアナトリアに移住したルートには2つの可能性がある
:東のカフカス山脈を越えた可能性、および西のバルカン半島を経た可能性である。
マロリー[2]とスタイナー[3]によればバルカン説の方がやや優勢である。
彼らがアナトリアに入ったのは紀元前2千年頃、またはそれ以前といわれる。
ただしコリン・レンフリューにより、印欧祖語の源郷をアナトリアとする説も提唱されている。
この説をとれば移住を仮定する必要はない。
その他アルメニアを源郷とする説をとれば、すぐ東から移動しただけということになる。
http://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%8A%E3%83%88%E3%83%AA%E3%82%A2%E8%AA%9E%E6%B4%BE
アナトリア語派(アナトリアごは、Anatolian languages)とは古代小アジア(アナトリア:現在のトルコ)で話されていたインド・ヨーロッパ語族の言語群で、
すべて死語である。中でもヒッタイト語の資料が多く最もよく研究されている。
その周辺で使われた言語もいくつか知られるが、これらについては資料が少ない。
起源
アナトリア語派はインド・ヨーロッパ祖語(印欧祖語)から最も早期に分かれたとの説が有力で、
一部では「インド・ヒッタイト語族」という言い方もされる。
この時期については紀元前4千年紀半ばとするクルガン仮説があるが、さらに古いとする説(レンフリュー説、また語彙統計学による報告[1])もある。
クルガン仮説によれば、初期のアナトリア語派話者が黒海の北からアナトリアに移住したルートには2つの可能性がある
:東のカフカス山脈を越えた可能性、および西のバルカン半島を経た可能性である。
マロリー[2]とスタイナー[3]によればバルカン説の方がやや優勢である。
彼らがアナトリアに入ったのは紀元前2千年頃、またはそれ以前といわれる。
ただしコリン・レンフリューにより、印欧祖語の源郷をアナトリアとする説も提唱されている。
この説をとれば移住を仮定する必要はない。
その他アルメニアを源郷とする説をとれば、すぐ東から移動しただけということになる。
>>929
言語[編集]
ヒッタイト語(nesili):紀元前1600年から1100年の頃使われたヒッタイト帝国の公用語。ヒッタイトの先住民が話していた原ハッティ語は、別系統である。Indo-Hittiteを参照。
ルウィ語(luwili):ヒッタイト語に近く、ヒッタイト帝国内でヒッタイト語に隣接する地域で話された。
楔形文字ルウィ語(英語版):ヒッタイト文書の中に楔形文字で書かれている。単語・短文のみ。
象形文字ルウィ語(英語版):印章・碑文に(アナトリア・ヒエログリフ(英語版))で書かれている。
リュキア語(英語版)(リュキア語Aまたは標準リュキア語)鉄器時代のリュキアで話された、ルウィ語の子孫言語。断片的。紀元前1世紀頃消えた。
ミリヤ語(英語版)(リュキア語B):リュキア語の方言で、碑文1つしか知られていない。
カリア語(英語版):カリアで話され、エジプトで見つかっている紀元前7世紀頃のカリア人傭兵の落書きで知られる。紀元前3世紀頃消えた。
ピシディア語(英語版)とシデ語(英語版)(パンフィリア語):断片的。
パラ語(英語版):アナトリア中北部で話され、紀元前13世紀頃消えた。ヒッタイト文書の中に祭文が引用されるのみ。
リュディア語(英語版):リュディアで話され、紀元前1世紀頃消えた。断片的。
他に、文字は残さなかったが、ミシア語(英語版)、カッパドキアやパンフィリアやパフラゴニア(英語版)にも同じ語派の言語があったようである。
言語[編集]
ヒッタイト語(nesili):紀元前1600年から1100年の頃使われたヒッタイト帝国の公用語。ヒッタイトの先住民が話していた原ハッティ語は、別系統である。Indo-Hittiteを参照。
ルウィ語(luwili):ヒッタイト語に近く、ヒッタイト帝国内でヒッタイト語に隣接する地域で話された。
楔形文字ルウィ語(英語版):ヒッタイト文書の中に楔形文字で書かれている。単語・短文のみ。
象形文字ルウィ語(英語版):印章・碑文に(アナトリア・ヒエログリフ(英語版))で書かれている。
リュキア語(英語版)(リュキア語Aまたは標準リュキア語)鉄器時代のリュキアで話された、ルウィ語の子孫言語。断片的。紀元前1世紀頃消えた。
ミリヤ語(英語版)(リュキア語B):リュキア語の方言で、碑文1つしか知られていない。
カリア語(英語版):カリアで話され、エジプトで見つかっている紀元前7世紀頃のカリア人傭兵の落書きで知られる。紀元前3世紀頃消えた。
ピシディア語(英語版)とシデ語(英語版)(パンフィリア語):断片的。
パラ語(英語版):アナトリア中北部で話され、紀元前13世紀頃消えた。ヒッタイト文書の中に祭文が引用されるのみ。
リュディア語(英語版):リュディアで話され、紀元前1世紀頃消えた。断片的。
他に、文字は残さなかったが、ミシア語(英語版)、カッパドキアやパンフィリアやパフラゴニア(英語版)にも同じ語派の言語があったようである。
http://web.kyoto-inet.or.jp/people/tiakio/antiGM/lat.html
ラト(Lat)
イタリア語のlatte(「乳」)は、乳を与えてくれる女神ラトから派生した。女神ラトは、ローマ以前の部族国家ラティウムの名祖だった。
この女神は、ラトナあるいはレートーとも呼ばれ、「世界卵」や太陽の母親だった[1]。ラトは、アラビア人たちにはアラート(「月」)の名で知られていた。
アラートは、のちに男神に変えられ、イスラム教のアラーになった。ラトは先王朝時代のエジプトでもよく知られており、
ヘーロドトスは彼女のことを「下エジプト王国の古代の女王」と呼んだ。
ラトの礼拝堂は、エジプト最古の神託所で、ギリシア人にはエスナ〔ラトポリス〕の名で知られていた[2]。
この神託所は、エジプト人にとっては、メンヘト(「月の家」)だった[3]。
ラトは、「月」だった。古代世界においては、月が万物の栄養源、すなわち、「乳の道」(天の川)Milky Wayを生み出した「天界の乳房」とみなされていたからである。
水、乳、血、樹液、ならびに生命を維持してくれるそのほかさまざまな液体は、すべて月から生じた。
ラトは、古代ラティウムにおける母権的文化の創始者だった。
latifundiaという古語は、女神ラトが自らの手で区画し、家母長たちに分配したそれぞれの土地という意味だった[4]。
Matrilineal Inheritance. マルタ島は、かつては、マー・ラタ(「母神ラト」)だった。
1428年にキリスト教会は、住民が異端思想をいだいている疑いがあるという理由で、
イタリアにある町を完全に破壊してしまったが、奇しくもその町の名は、マグナラタ(「大いなるラト」)だった。
[1]Graves, W. G., 318.
[2]Larousse, 29.
[3]Budge, G. E. 2, 50.
[4]Cumont, M. M., 74.
Barbara G. Walker : The Woman's Encyclopedia of Myths and Secrets (Harper & Row, 1983)
ラト(Lat)
イタリア語のlatte(「乳」)は、乳を与えてくれる女神ラトから派生した。女神ラトは、ローマ以前の部族国家ラティウムの名祖だった。
この女神は、ラトナあるいはレートーとも呼ばれ、「世界卵」や太陽の母親だった[1]。ラトは、アラビア人たちにはアラート(「月」)の名で知られていた。
アラートは、のちに男神に変えられ、イスラム教のアラーになった。ラトは先王朝時代のエジプトでもよく知られており、
ヘーロドトスは彼女のことを「下エジプト王国の古代の女王」と呼んだ。
ラトの礼拝堂は、エジプト最古の神託所で、ギリシア人にはエスナ〔ラトポリス〕の名で知られていた[2]。
この神託所は、エジプト人にとっては、メンヘト(「月の家」)だった[3]。
ラトは、「月」だった。古代世界においては、月が万物の栄養源、すなわち、「乳の道」(天の川)Milky Wayを生み出した「天界の乳房」とみなされていたからである。
水、乳、血、樹液、ならびに生命を維持してくれるそのほかさまざまな液体は、すべて月から生じた。
ラトは、古代ラティウムにおける母権的文化の創始者だった。
latifundiaという古語は、女神ラトが自らの手で区画し、家母長たちに分配したそれぞれの土地という意味だった[4]。
Matrilineal Inheritance. マルタ島は、かつては、マー・ラタ(「母神ラト」)だった。
1428年にキリスト教会は、住民が異端思想をいだいている疑いがあるという理由で、
イタリアにある町を完全に破壊してしまったが、奇しくもその町の名は、マグナラタ(「大いなるラト」)だった。
[1]Graves, W. G., 318.
[2]Larousse, 29.
[3]Budge, G. E. 2, 50.
[4]Cumont, M. M., 74.
Barbara G. Walker : The Woman's Encyclopedia of Myths and Secrets (Harper & Row, 1983)
このネタは入ってなかった気がするな
まぁ、テンプル騎士団の方がメインだからね。
後、年代が合わない。>>931
http://www.coara.or.jp/~sanken/beppudic/preedo.html
1315
*教皇マルティヌス5世は清貧の思想を受け継いだフラティチェリ派(後の「フランシスコ会精神派」)を破門し、
フランティチェリ派のすみかであるマグナラタの村を襲撃して住民を皆殺しせよと命じた。
キエティスム
http://ja.wikipedia.org/wiki/%E3%82%AD%E3%82%A8%E3%83%86%E3%82%A3%E3%82%B9%E3%83%A0
キエティスム(英語読みは、クワイエティズム)(Quietism、静寂主義) は、様々な意味と定義を持つ用語である。
フラティチェリ(Fraticelli)の側では個人の自律に関する同様の主張が、1317年、ローマ教皇ヨハネス22世によって激しい糾弾にさらされた。
1329年、同教皇はマイスター・エックハルトの誤りの中にある、パンがキリストの肉体に変わったという秘蹟(→「化体説」の項を参照)と同様に、
我々は総合的に神へと変容するのだという主張、および、内なる行動の価値―これらは持続的に我々に与えられている神格によって造られたとされる―を法的に禁じた。
まぁ、テンプル騎士団の方がメインだからね。
後、年代が合わない。>>931
http://www.coara.or.jp/~sanken/beppudic/preedo.html
1315
*教皇マルティヌス5世は清貧の思想を受け継いだフラティチェリ派(後の「フランシスコ会精神派」)を破門し、
フランティチェリ派のすみかであるマグナラタの村を襲撃して住民を皆殺しせよと命じた。
キエティスム
http://ja.wikipedia.org/wiki/%E3%82%AD%E3%82%A8%E3%83%86%E3%82%A3%E3%82%B9%E3%83%A0
キエティスム(英語読みは、クワイエティズム)(Quietism、静寂主義) は、様々な意味と定義を持つ用語である。
フラティチェリ(Fraticelli)の側では個人の自律に関する同様の主張が、1317年、ローマ教皇ヨハネス22世によって激しい糾弾にさらされた。
1329年、同教皇はマイスター・エックハルトの誤りの中にある、パンがキリストの肉体に変わったという秘蹟(→「化体説」の項を参照)と同様に、
我々は総合的に神へと変容するのだという主張、および、内なる行動の価値―これらは持続的に我々に与えられている神格によって造られたとされる―を法的に禁じた。
>>931
シャーラダー文字
http://www.chikyukotobamura.org/muse/wr_sasia_15.html
シャーラダーという名称は,カシュミールで信仰されていた女神シャーラーダー・デーヴィー(??rad? Dev?)による。
シャーラダーとはシヴァ神の妃パーラヴァティーのことで,カシュミールの住民は,シャクティ(?akti)女神の信奉者(??kta)であったことから,この名称がついた。
??rad? script
http://en.wikipedia.org/wiki/%C5%9A%C4%81rad%C4%81_script
アナーヒター
http://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%8A%E3%83%BC%E3%83%92%E3%82%BF%E3%83%BC
アナーヒターはアヴェスター語形で、パフラヴィー語ではアナーヒト(An?hit)といい、いずれも「清浄」を意味する。他にもナーキッドという別名もある。
姿形[編集]
アナーヒターは力強い色白の腕を持ち、四角い黄金の耳飾りと、百の星をちりばめた黄金の冠をかぶり、
黄金のマントを羽織り、首には黄金の首飾りを身に付け、帯を高く締めた美しい乙女の姿をしているといわれる。
バビロニアのイシュタルと習合し、このとき容貌、装束、性格、祭儀もバビロニア的となる。
ヘレニズム時代になるとギリシャでも崇められるようになり、アフロディーテとも習合した。
ギリシャ語でアナイティス(’Ανα?^τι?)と呼ばれる。
また、リュディアではキュベレやアルテミスと同一視された。
シャーラダー文字
http://www.chikyukotobamura.org/muse/wr_sasia_15.html
シャーラダーという名称は,カシュミールで信仰されていた女神シャーラーダー・デーヴィー(??rad? Dev?)による。
シャーラダーとはシヴァ神の妃パーラヴァティーのことで,カシュミールの住民は,シャクティ(?akti)女神の信奉者(??kta)であったことから,この名称がついた。
??rad? script
http://en.wikipedia.org/wiki/%C5%9A%C4%81rad%C4%81_script
アナーヒター
http://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%8A%E3%83%BC%E3%83%92%E3%82%BF%E3%83%BC
アナーヒターはアヴェスター語形で、パフラヴィー語ではアナーヒト(An?hit)といい、いずれも「清浄」を意味する。他にもナーキッドという別名もある。
姿形[編集]
アナーヒターは力強い色白の腕を持ち、四角い黄金の耳飾りと、百の星をちりばめた黄金の冠をかぶり、
黄金のマントを羽織り、首には黄金の首飾りを身に付け、帯を高く締めた美しい乙女の姿をしているといわれる。
バビロニアのイシュタルと習合し、このとき容貌、装束、性格、祭儀もバビロニア的となる。
ヘレニズム時代になるとギリシャでも崇められるようになり、アフロディーテとも習合した。
ギリシャ語でアナイティス(’Ανα?^τι?)と呼ばれる。
また、リュディアではキュベレやアルテミスと同一視された。
知ってるのはこっち
>>931
http://ja.wikipedia.org/wiki/%E5%A4%A9%E3%81%AE%E5%B7%9D
ギリシャ神話[編集]ギリシャ語では夜空の光の帯を「γαλαξ?α?(galaxias)」もしくは「kyklos galaktikos」と言う。kyklos galaktikosは「乳の環[5]」という意味。
それにまつわるギリシャ神話を紹介する。
ゼウスは、自分とアルクメネの子のヘラクレスを不死身にするために、女神ヘラの母乳をヘラクレスに飲ませようとしていた。
しかし、嫉妬深いヘラはヘラクレスを憎んでいたため母乳を飲ませようとはしなかった。
一計を案じたゼウスはヘラに眠り薬を飲ませ、ヘラが眠っているあいだにヘラクレスに母乳を飲ませた。
この時、ヘラが目覚め、ヘラクレスが自分の乳を飲んでいることに驚き、払いのけた際にヘラの母乳が流れ出した。これが天のミルクの環になった。
ヘーラー
http://ja.wikipedia.org/wiki/%E3%83%98%E3%83%BC%E3%83%A9%E3%83%BC
ヘーラーの母乳は飲んだ人間の肉体を強化し不死身にする力があり、ヘーラクレースもこれを飲んだため乳児時代から驚異的な怪力を発揮することが出来た。
また、この時へーラクレースの母乳を吸う力があまりにも強かった為、へーラーはヘーラクレースを突き飛ばし、その際に飛び散ったへーラーの母乳が天の川になった。
なお、ヘーラクレースは、ヘーラーの子ではないが「ヘーラーの栄光」と言う意味の名を持つ[1]。
崇拝[編集]
ヘーラーはサモス島で誕生したと考えられており、サモス島は古くからヘーラー信仰の中心地となっていた。
元来は、アルゴス、ミュケーナイ、スパルタ等のペロポネーソス半島一帯に確固たる宗教的基盤を持っており、
かつてアカイア人に信仰された地母神であったとされ、北方からの征服者との和合をゼウスとの結婚で象徴させたと考えられる[2]。
二神の不和は、両者の崇拝者が敵対関係にあったことの名残とも考えられている[2]。
>>931
http://ja.wikipedia.org/wiki/%E5%A4%A9%E3%81%AE%E5%B7%9D
ギリシャ神話[編集]ギリシャ語では夜空の光の帯を「γαλαξ?α?(galaxias)」もしくは「kyklos galaktikos」と言う。kyklos galaktikosは「乳の環[5]」という意味。
それにまつわるギリシャ神話を紹介する。
ゼウスは、自分とアルクメネの子のヘラクレスを不死身にするために、女神ヘラの母乳をヘラクレスに飲ませようとしていた。
しかし、嫉妬深いヘラはヘラクレスを憎んでいたため母乳を飲ませようとはしなかった。
一計を案じたゼウスはヘラに眠り薬を飲ませ、ヘラが眠っているあいだにヘラクレスに母乳を飲ませた。
この時、ヘラが目覚め、ヘラクレスが自分の乳を飲んでいることに驚き、払いのけた際にヘラの母乳が流れ出した。これが天のミルクの環になった。
ヘーラー
http://ja.wikipedia.org/wiki/%E3%83%98%E3%83%BC%E3%83%A9%E3%83%BC
ヘーラーの母乳は飲んだ人間の肉体を強化し不死身にする力があり、ヘーラクレースもこれを飲んだため乳児時代から驚異的な怪力を発揮することが出来た。
また、この時へーラクレースの母乳を吸う力があまりにも強かった為、へーラーはヘーラクレースを突き飛ばし、その際に飛び散ったへーラーの母乳が天の川になった。
なお、ヘーラクレースは、ヘーラーの子ではないが「ヘーラーの栄光」と言う意味の名を持つ[1]。
崇拝[編集]
ヘーラーはサモス島で誕生したと考えられており、サモス島は古くからヘーラー信仰の中心地となっていた。
元来は、アルゴス、ミュケーナイ、スパルタ等のペロポネーソス半島一帯に確固たる宗教的基盤を持っており、
かつてアカイア人に信仰された地母神であったとされ、北方からの征服者との和合をゼウスとの結婚で象徴させたと考えられる[2]。
二神の不和は、両者の崇拝者が敵対関係にあったことの名残とも考えられている[2]。
>>932
ヨハネス22世 (ローマ教皇)
http://ja.wikipedia.org/wiki/%E3%83%A8%E3%83%8F%E3%83%8D%E3%82%B922%E4%B8%96_(%E3%83%AD%E3%83%BC%E3%83%9E%E6%95%99%E7%9A%87)
1309年に教皇庁が南仏アヴィニョンに移るという事態が生じた(「アヴィニョン捕囚」)。
アヴィニョン教皇庁での初めての教皇となったクレメンス5世は、修道会の会則を厳格に遵守するかどうか(「清貧論争」))をめぐって
分裂傾向にあったフランシスコ会(フランチェスコ会)の問題を調査する委員会を教皇庁内に設けた[3]。
http://en.wikipedia.org/wiki/Fraticelli
>>932
Early in 1317 John XXII, pursuant to a decree of Boniface VIII, declared Angelo excommunicated and placed him in custody.
He defended himself ably in his "Epistola Excusatoria", representing himself as a zealous Franciscan, but John XXII refused to admit his plea,
Angelo being a Celestine hermit, and in the decree "Sancta Romana et universalis ecclesia" (30 December 1317) refused to authorize the congregation of which Angelo was head.
>>931
On 27 January of the same year,
Martin V permitted the Observants of Ancona to occupy the monastery of the Fraticelli at Castro l'Ermita as a first step in the campaign against the Fraticelli of that neighbourhood.
On 1 June 1428, he commanded the Bishop of Ancona to enforce his rulings strictly in Maiolati,
to put all suspects to the rack, destroy their village, separate the children from heretical parents, and disperse the elder population.
A circular letter, which the Fraticelli addressed to all Christendom, proved ineffectual and their doom was sealed.
John of Capistrano and James of the March burned thirty-six of their establishments or dispersed the members and a number were burned at the stake at Florence and Fabriano,
at the latter place in the presence of the pope.
ヨハネス22世 (ローマ教皇)
http://ja.wikipedia.org/wiki/%E3%83%A8%E3%83%8F%E3%83%8D%E3%82%B922%E4%B8%96_(%E3%83%AD%E3%83%BC%E3%83%9E%E6%95%99%E7%9A%87)
1309年に教皇庁が南仏アヴィニョンに移るという事態が生じた(「アヴィニョン捕囚」)。
アヴィニョン教皇庁での初めての教皇となったクレメンス5世は、修道会の会則を厳格に遵守するかどうか(「清貧論争」))をめぐって
分裂傾向にあったフランシスコ会(フランチェスコ会)の問題を調査する委員会を教皇庁内に設けた[3]。
http://en.wikipedia.org/wiki/Fraticelli
>>932
Early in 1317 John XXII, pursuant to a decree of Boniface VIII, declared Angelo excommunicated and placed him in custody.
He defended himself ably in his "Epistola Excusatoria", representing himself as a zealous Franciscan, but John XXII refused to admit his plea,
Angelo being a Celestine hermit, and in the decree "Sancta Romana et universalis ecclesia" (30 December 1317) refused to authorize the congregation of which Angelo was head.
>>931
On 27 January of the same year,
Martin V permitted the Observants of Ancona to occupy the monastery of the Fraticelli at Castro l'Ermita as a first step in the campaign against the Fraticelli of that neighbourhood.
On 1 June 1428, he commanded the Bishop of Ancona to enforce his rulings strictly in Maiolati,
to put all suspects to the rack, destroy their village, separate the children from heretical parents, and disperse the elder population.
A circular letter, which the Fraticelli addressed to all Christendom, proved ineffectual and their doom was sealed.
John of Capistrano and James of the March burned thirty-six of their establishments or dispersed the members and a number were burned at the stake at Florence and Fabriano,
at the latter place in the presence of the pope.
>>801
馬の家畜化と搾乳は5500年前から
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=63936611
乗馬とウマの搾乳の先駆者は、いまはカザフスタンの国土となっている広大な草原に暮らしていた人々だった。
少なくとも5500年前、この地域の人々がウマを家畜として飼い慣らしていたという新しい証拠が見つかった。
「ウマを食用としただけでなく、その乳も利用していた。この地域では、非常に早い段階から乗馬と搾乳の両方が行われていた」と、
今回の研究を指揮したイギリス、エクセター大学の考古学者アラン・ウートラム氏は話す。
この発見により、ウマの家畜化の開始時期は1000年早まることになる。
ウマがいつ頃から家畜化されていたかについては長年の研究対象だった。(略)
以前の研究では、3600〜3100年前にカザフスタン北部に生きていたボタイ人の間で、ウマが家畜化されていたことを示す状況証拠が見つかっていた。
馬具の作製に使用したと考えられる革細工道具が発見されていたのである。
今回の研究では、過去数年間にボタイ遺跡で出土した骨や陶器の調査が行われた。
「ボタイでウマが家畜化されていたことを示す、まったく異なる3つの証拠を入手できた。
どれも強力な証拠だ」とウートラム氏は言う。同氏の研究成果は、3月5日発行の「Science」誌で発表された。
1つ目の証拠は、歯の摩耗(まもう)である。出土したウマの骨を調べたところ、数頭のウマの歯にはハミ(馬銜、銜)という馬具を噛んでいたと考えられる摩耗があった。
研究チームは、すり切れた歯の1つは紀元前3500年頃のものと推定している。
2つ目は野生種と比べて細い脚の骨で、これは家畜化されているウマの特徴である。
3つ目の証拠は、陶器のつぼの縁辺りに相当する破片から検出されたウマの脂肪と馬乳の脂肪分だ。
ニューヨークにあるハートウィック大学の考古学者デイビッド・アンソニー氏に専門家の意見を聞いたところ、
「土器に付着した脂肪で、ウマの脂肪組織だけでなく、馬乳の脂肪も特定できたことは非常に重要なことだ。
野生のウマから搾乳はしない」と述べている。
以上の証拠から、人類はウマを家畜化してまもなくその乳の利用を思い付いたらしい。
馬の家畜化と搾乳は5500年前から
http://www.nationalgeographic.co.jp/news/news_article.php?file_id=63936611
乗馬とウマの搾乳の先駆者は、いまはカザフスタンの国土となっている広大な草原に暮らしていた人々だった。
少なくとも5500年前、この地域の人々がウマを家畜として飼い慣らしていたという新しい証拠が見つかった。
「ウマを食用としただけでなく、その乳も利用していた。この地域では、非常に早い段階から乗馬と搾乳の両方が行われていた」と、
今回の研究を指揮したイギリス、エクセター大学の考古学者アラン・ウートラム氏は話す。
この発見により、ウマの家畜化の開始時期は1000年早まることになる。
ウマがいつ頃から家畜化されていたかについては長年の研究対象だった。(略)
以前の研究では、3600〜3100年前にカザフスタン北部に生きていたボタイ人の間で、ウマが家畜化されていたことを示す状況証拠が見つかっていた。
馬具の作製に使用したと考えられる革細工道具が発見されていたのである。
今回の研究では、過去数年間にボタイ遺跡で出土した骨や陶器の調査が行われた。
「ボタイでウマが家畜化されていたことを示す、まったく異なる3つの証拠を入手できた。
どれも強力な証拠だ」とウートラム氏は言う。同氏の研究成果は、3月5日発行の「Science」誌で発表された。
1つ目の証拠は、歯の摩耗(まもう)である。出土したウマの骨を調べたところ、数頭のウマの歯にはハミ(馬銜、銜)という馬具を噛んでいたと考えられる摩耗があった。
研究チームは、すり切れた歯の1つは紀元前3500年頃のものと推定している。
2つ目は野生種と比べて細い脚の骨で、これは家畜化されているウマの特徴である。
3つ目の証拠は、陶器のつぼの縁辺りに相当する破片から検出されたウマの脂肪と馬乳の脂肪分だ。
ニューヨークにあるハートウィック大学の考古学者デイビッド・アンソニー氏に専門家の意見を聞いたところ、
「土器に付着した脂肪で、ウマの脂肪組織だけでなく、馬乳の脂肪も特定できたことは非常に重要なことだ。
野生のウマから搾乳はしない」と述べている。
以上の証拠から、人類はウマを家畜化してまもなくその乳の利用を思い付いたらしい。
>>801
デレイフカ遺跡とボタイ遺跡/馬の騎乗はいつからか?
http://www.rui.jp/ruinet.html?i=200&c=400&m=246216
(から抜粋)
「スキタイと匈奴 遊牧の文明」林俊雄 講談社より、関連箇所の要約を以下にまとめます。
同様に馬の骨が大量に発掘された遺跡に、カザフスタン北部のボタイ遺跡がある。
ボタイ遺跡は銅石器時代の集落址で、1970年代から1980年代にかけての15シーズンにわたる調査で30万点(10万トン)にのぼる骨の資料が出土し、そのうち99.9パーセントが馬の骨であった。
(この遺跡の)補正された年代は、5500〜5000年前とされている。
デレイフカの年代についても、黒海北岸の他の文化との比較から5700〜5150年前とし、ボタイとほぼ重なるとも付け加えている。
その後馬の骨の放射性炭素による年代測定で、馬の頭骨は前790〜前520年、歯は前410〜前200年とわかり、
この馬に関しては後世のスキタイ時代の埋納と訂正せざるを得なくなってしまった。
また、「銜(はみ)留め具」と称する骨角製品に関しても、ボタイの骨製品は馬の革帯を通すには孔が小さすぎ、
また孔が一つしかない製品では、銜(はみ)留め具としての役割を果たすことができないことから、6000〜5000年に馬の騎乗はなかったと一般には結論づけらる。
デレイフカ遺跡とボタイ遺跡/馬の騎乗はいつからか?
http://www.rui.jp/ruinet.html?i=200&c=400&m=246216
(から抜粋)
「スキタイと匈奴 遊牧の文明」林俊雄 講談社より、関連箇所の要約を以下にまとめます。
同様に馬の骨が大量に発掘された遺跡に、カザフスタン北部のボタイ遺跡がある。
ボタイ遺跡は銅石器時代の集落址で、1970年代から1980年代にかけての15シーズンにわたる調査で30万点(10万トン)にのぼる骨の資料が出土し、そのうち99.9パーセントが馬の骨であった。
(この遺跡の)補正された年代は、5500〜5000年前とされている。
デレイフカの年代についても、黒海北岸の他の文化との比較から5700〜5150年前とし、ボタイとほぼ重なるとも付け加えている。
その後馬の骨の放射性炭素による年代測定で、馬の頭骨は前790〜前520年、歯は前410〜前200年とわかり、
この馬に関しては後世のスキタイ時代の埋納と訂正せざるを得なくなってしまった。
また、「銜(はみ)留め具」と称する骨角製品に関しても、ボタイの骨製品は馬の革帯を通すには孔が小さすぎ、
また孔が一つしかない製品では、銜(はみ)留め具としての役割を果たすことができないことから、6000〜5000年に馬の騎乗はなかったと一般には結論づけらる。
>>806
http://en.wikipedia.org/wiki/Fjord_horse
History[edit]
Gloppen coat of armsThe Fjord horse is one of the world's oldest and purest breeds.
Horse were known to exist in Norway at the end of the last ice age.
It is believed that the ancestors of the modern Fjord horse migrated to Norway and were domesticated over 4,000 years ago.
Archaeological excavations at Viking burial sites indicate that the Fjord horse type has been selectively bred for at least 2,000 years.
http://en.wikipedia.org/wiki/Fjord_horse
History[edit]
Gloppen coat of armsThe Fjord horse is one of the world's oldest and purest breeds.
Horse were known to exist in Norway at the end of the last ice age.
It is believed that the ancestors of the modern Fjord horse migrated to Norway and were domesticated over 4,000 years ago.
Archaeological excavations at Viking burial sites indicate that the Fjord horse type has been selectively bred for at least 2,000 years.
>>926
http://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%95%E3%82%A1%E3%83%8A%E3%82%B7%E3%82%A7%E3%83%B4%E3%82%A9%E6%96%87%E5%8C%96
>>925
http://en.wikipedia.org/wiki/Tocharians
Tarim mummies[edit]
The Tarim Basin mummies (1800 BC) have been found in the same general geographical area as the Tocharian texts and frescoes from the Tarim Basin (3rd to 9th centuries AD),
and are both connected to an Indo-European origin and point to Caucasoid types with light eyes and hair color.
However it is unknown whether the mummies and frescoes are connected.
In 2008, the remains of another male were discovered near Turpan.
Thought by researchers to be a member of the Gushi culture, the man was buried with a number of practical and ceremonial objects,
including archery equipment and a harp, and 789 grams of cannabis.
Through genetic analysis and carbon dating, the burial has been dated to roughly 700 BC.
Only two of the 500 graves at the site contain cannabis, leading researchers to suggest shamanic roles for the two individuals.
In 2009, the remains of 30 individuals found at the Xiaohe Tomb complex were analyzed for Y-DNA and mtDNA markers.
They suggest that an admixed population of both western and eastern origin lived in the Tarim basin since the early Bronze Age.
The Xiaohe people maternal lineages were predominantly East Asian haplogroup C with smaller numbers of H and K, while the paternal lines were all West Eurasian R1a1a.
The geographic location of this admixing is unknown, although south Siberia is likely.[9]
http://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%95%E3%82%A1%E3%83%8A%E3%82%B7%E3%82%A7%E3%83%B4%E3%82%A9%E6%96%87%E5%8C%96
>>925
http://en.wikipedia.org/wiki/Tocharians
Tarim mummies[edit]
The Tarim Basin mummies (1800 BC) have been found in the same general geographical area as the Tocharian texts and frescoes from the Tarim Basin (3rd to 9th centuries AD),
and are both connected to an Indo-European origin and point to Caucasoid types with light eyes and hair color.
However it is unknown whether the mummies and frescoes are connected.
In 2008, the remains of another male were discovered near Turpan.
Thought by researchers to be a member of the Gushi culture, the man was buried with a number of practical and ceremonial objects,
including archery equipment and a harp, and 789 grams of cannabis.
Through genetic analysis and carbon dating, the burial has been dated to roughly 700 BC.
Only two of the 500 graves at the site contain cannabis, leading researchers to suggest shamanic roles for the two individuals.
In 2009, the remains of 30 individuals found at the Xiaohe Tomb complex were analyzed for Y-DNA and mtDNA markers.
They suggest that an admixed population of both western and eastern origin lived in the Tarim basin since the early Bronze Age.
The Xiaohe people maternal lineages were predominantly East Asian haplogroup C with smaller numbers of H and K, while the paternal lines were all West Eurasian R1a1a.
The geographic location of this admixing is unknown, although south Siberia is likely.[9]
>>939
Evidence that a West-East admixed population lived in the Tarim Basin as early as the early Bronze Age
http://www.biomedcentral.com/1741-7007/8/15#IDAH0OBH
DNA amplification, cloning, and sequencing
The nucleotide positions 16035-16409 of the mitochondrial genome was amplified by two overlapping primer pairs.
In addition, a number of coding-region mtDNA polymorphisms were typed, which are diagnostic for major branches in the
mtDNA tree: Haplogroups R(12705C), UK(12308G), HV(14766T), H(7028C), R1(4917G), R11(10031C) and C4(11969A) were identified by direct sequencing
and haplogroups M(10400T), F(3970T) and C(14318C) were examined by using amplified product-length polymorphisms method [11-13].
The B haplogroup was identified based on 9-bp deletion in np8280.
Some Y-chromosomal single nucleotide polymorphisms (Y-SNPs) were typed, which are diagnostic for major branches in the Y chromosome haplogroup tree [14,15]:
Haplogroups F(M89T), K(M9G), P(M45A), R1(M173A), and R1a1a(M198A) were identified by direct sequencing.
The primers used in HVRI and diagnostic SNP markers are shown in Table 2.
Phylogenetic tree of haplogroup C, based on HVS-I sequences in the region 16050-16391
http://www.biomedcentral.com/1741-7007/8/15/figure/F4
Map of Eurasia, showing ancient populations from the Tarim Basin and surroundings
http://www.biomedcentral.com/1741-7007/8/15/figure/F5
うーんこれどうなんかな
Evidence that a West-East admixed population lived in the Tarim Basin as early as the early Bronze Age
http://www.biomedcentral.com/1741-7007/8/15#IDAH0OBH
DNA amplification, cloning, and sequencing
The nucleotide positions 16035-16409 of the mitochondrial genome was amplified by two overlapping primer pairs.
In addition, a number of coding-region mtDNA polymorphisms were typed, which are diagnostic for major branches in the
mtDNA tree: Haplogroups R(12705C), UK(12308G), HV(14766T), H(7028C), R1(4917G), R11(10031C) and C4(11969A) were identified by direct sequencing
and haplogroups M(10400T), F(3970T) and C(14318C) were examined by using amplified product-length polymorphisms method [11-13].
The B haplogroup was identified based on 9-bp deletion in np8280.
Some Y-chromosomal single nucleotide polymorphisms (Y-SNPs) were typed, which are diagnostic for major branches in the Y chromosome haplogroup tree [14,15]:
Haplogroups F(M89T), K(M9G), P(M45A), R1(M173A), and R1a1a(M198A) were identified by direct sequencing.
The primers used in HVRI and diagnostic SNP markers are shown in Table 2.
Phylogenetic tree of haplogroup C, based on HVS-I sequences in the region 16050-16391
http://www.biomedcentral.com/1741-7007/8/15/figure/F4
Map of Eurasia, showing ancient populations from the Tarim Basin and surroundings
http://www.biomedcentral.com/1741-7007/8/15/figure/F5
うーんこれどうなんかな
941 :世界@名無史さん:2014/02/10(月) 20:07:49.84 0
時代が下るほど東ユーラシア人の血統が濃くなる感じかな。
まあ、地理的な位置を考えれば当然か。
まあ、地理的な位置を考えれば当然か。
>>926>>940
コーカソイド
>>559>>703
http://2.bp.blogspot.com/-rhqReVApmRk/Un1LdnrSSyI/AAAAAAAAJY8/cmRVUA1On9k/s1600/journal.pgen.1003912.g002.png
タジキスタンからパキスタンの辺りでタリム盆地側では無さそう
ただミヌシンスク盆地側にはもうちょっと伸びてる
コーカソイド
>>559>>703
http://2.bp.blogspot.com/-rhqReVApmRk/Un1LdnrSSyI/AAAAAAAAJY8/cmRVUA1On9k/s1600/journal.pgen.1003912.g002.png
タジキスタンからパキスタンの辺りでタリム盆地側では無さそう
ただミヌシンスク盆地側にはもうちょっと伸びてる
ソグディアナ
http://ja.wikipedia.org/wiki/%E3%82%BD%E3%82%B0%E3%83%87%E3%82%A3%E3%82%A2%E3%83%8A
ソグディアナ(Sogdiana)は、中央アジアのアムダリヤ川とシルダリヤ川の中間に位置し、サマルカンドを中心的な都市とするザラフシャン川流域地方の古名。
バクトリアの北、ホラズムの東、康居の南東に位置する地方。
現在のウズベキスタンのサマルカンド州とブハラ州、タジキスタンのソグド州に相当する。
イラン系の言語を話すソグド人と呼ばれる民族が居住したことからつけられたギリシャ語・ラテン語の名称で、
西からみてオクサス川(アムダリヤ川)の向こう側にあることから、トランスオクシアナとも呼ばれる。
中国の歴史書では粟特と記されている。
ソグド語
http://ja.wikipedia.org/wiki/%E3%82%BD%E3%82%B0%E3%83%89%E8%AA%9E
ソグド語は、中世ペルシャ語(英語版)やパルティア語(英語版)と並んで、中期イラン諸語の中でも重要な言語のひとつであり、多くの言語資料が遺されている。
ソグド語は文法や形態論は中期ペルシア語と比べてより伝統的な形である。
ソグド語はイラン語派の北東語群に属する。ソグド語は中央アジア(トランスオクシアナ)における交易言語であり、中国人商人とイラン人商人との間の共通語でもあった。
ソグド語は経済的・政治的に重要な言語であったため、8世紀初頭にソグディアナがムスリムによって征服された後も数世紀にわたって使用され続けた。
表記体系[編集]
初期のソグド文字の例はソグディアナ地域において発見されている。
これらはアラム文字を元にしたものであり、表記と発音が大きく異なっていた。
ソグド文字からはウイグル文字が派生し、ウイグル文字は後にモンゴル文字の元となった。
法隆寺で見つかった650年頃のものと思われる香木に、ソグド文字が書かれていた例がある[1]。
http://ja.wikipedia.org/wiki/%E3%82%BD%E3%82%B0%E3%83%87%E3%82%A3%E3%82%A2%E3%83%8A
ソグディアナ(Sogdiana)は、中央アジアのアムダリヤ川とシルダリヤ川の中間に位置し、サマルカンドを中心的な都市とするザラフシャン川流域地方の古名。
バクトリアの北、ホラズムの東、康居の南東に位置する地方。
現在のウズベキスタンのサマルカンド州とブハラ州、タジキスタンのソグド州に相当する。
イラン系の言語を話すソグド人と呼ばれる民族が居住したことからつけられたギリシャ語・ラテン語の名称で、
西からみてオクサス川(アムダリヤ川)の向こう側にあることから、トランスオクシアナとも呼ばれる。
中国の歴史書では粟特と記されている。
ソグド語
http://ja.wikipedia.org/wiki/%E3%82%BD%E3%82%B0%E3%83%89%E8%AA%9E
ソグド語は、中世ペルシャ語(英語版)やパルティア語(英語版)と並んで、中期イラン諸語の中でも重要な言語のひとつであり、多くの言語資料が遺されている。
ソグド語は文法や形態論は中期ペルシア語と比べてより伝統的な形である。
ソグド語はイラン語派の北東語群に属する。ソグド語は中央アジア(トランスオクシアナ)における交易言語であり、中国人商人とイラン人商人との間の共通語でもあった。
ソグド語は経済的・政治的に重要な言語であったため、8世紀初頭にソグディアナがムスリムによって征服された後も数世紀にわたって使用され続けた。
表記体系[編集]
初期のソグド文字の例はソグディアナ地域において発見されている。
これらはアラム文字を元にしたものであり、表記と発音が大きく異なっていた。
ソグド文字からはウイグル文字が派生し、ウイグル文字は後にモンゴル文字の元となった。
法隆寺で見つかった650年頃のものと思われる香木に、ソグド文字が書かれていた例がある[1]。
バクトリア
http://ja.wikipedia.org/wiki/%E3%83%90%E3%82%AF%E3%83%88%E3%83%AA%E3%82%A2
オアシス都市バークトリシュ
バクトリアがもっとも古く現れているのは、アケメネス朝のダレイオス1世(在位:前522年 - 前486年)によって建てられた『ベヒストゥン碑文』にバークトリシュ(Bakhtri?)とあるのが初見である。
バクトリアはヘラート、メルヴ、サマルカンドなどとともに中央アジアにおけるイラン系民族によって建設された最古のオアシス都市のひとつである。
首都のバクトラは現在のバルフに比定されている。
現在、バルフは人口数百の小さな村落にすぎないが、村の近くには城壁に囲まれた巨大な廃墟がある。
今までのところバクトラに比定すべき証拠は発見されていない。
一説には現在のバルフは古代のバクトラではなく、バクトラはもっとアム川に近いところにあったともいわれている。
しかし、バルフの付近には古代ゾロアスター教の祭壇の遺跡もあり、バクトリアの貨幣も多く出土している。
ゾロアスター教の中心地
ペルシア文明に大きな影響を与えたゾロアスター教の開祖であるゾロアスターは、古くからバクトリアの人だという伝説がある。
少なくともアケメネス朝時代にはバクトラがゾロアスター教の中心地の一つであったことは明らかである。
また、古いペルシアの伝説では、ゾロアスターはアレクサンドロスの侵入より258年前の人だとされている。
ペルシア人とサカ人の共同支配
紀元前6世紀のバクトリアの人口の主要部分、少なくともその支配階級はペルシア人とスキタイ人とによって構成されていたものと考えられるが、
スキタイ系のサカ人がこの地域にペルシア人よりも早くから住んでいたことは、ヘロドトスや、その後のストラボンなどが伝えているバクトリアの習俗によっても知られる。
サカ人に遅れてバクトリアに侵入し、これを征服したペルシア人は、サカ人やその他の先住民の支配者となったが、その数は多くはなかった。
ローマ時代の史料によると、バクトリアは7000人の貴族によって支配されていたという。
また、ヘロドトスがしばしば「バクトリア人とサカ人」と並称させているところからみても、ペルシア人と先住のサカ人が共同して支配階級を構成していたものと思われる。
http://ja.wikipedia.org/wiki/%E3%83%90%E3%82%AF%E3%83%88%E3%83%AA%E3%82%A2
オアシス都市バークトリシュ
バクトリアがもっとも古く現れているのは、アケメネス朝のダレイオス1世(在位:前522年 - 前486年)によって建てられた『ベヒストゥン碑文』にバークトリシュ(Bakhtri?)とあるのが初見である。
バクトリアはヘラート、メルヴ、サマルカンドなどとともに中央アジアにおけるイラン系民族によって建設された最古のオアシス都市のひとつである。
首都のバクトラは現在のバルフに比定されている。
現在、バルフは人口数百の小さな村落にすぎないが、村の近くには城壁に囲まれた巨大な廃墟がある。
今までのところバクトラに比定すべき証拠は発見されていない。
一説には現在のバルフは古代のバクトラではなく、バクトラはもっとアム川に近いところにあったともいわれている。
しかし、バルフの付近には古代ゾロアスター教の祭壇の遺跡もあり、バクトリアの貨幣も多く出土している。
ゾロアスター教の中心地
ペルシア文明に大きな影響を与えたゾロアスター教の開祖であるゾロアスターは、古くからバクトリアの人だという伝説がある。
少なくともアケメネス朝時代にはバクトラがゾロアスター教の中心地の一つであったことは明らかである。
また、古いペルシアの伝説では、ゾロアスターはアレクサンドロスの侵入より258年前の人だとされている。
ペルシア人とサカ人の共同支配
紀元前6世紀のバクトリアの人口の主要部分、少なくともその支配階級はペルシア人とスキタイ人とによって構成されていたものと考えられるが、
スキタイ系のサカ人がこの地域にペルシア人よりも早くから住んでいたことは、ヘロドトスや、その後のストラボンなどが伝えているバクトリアの習俗によっても知られる。
サカ人に遅れてバクトリアに侵入し、これを征服したペルシア人は、サカ人やその他の先住民の支配者となったが、その数は多くはなかった。
ローマ時代の史料によると、バクトリアは7000人の貴族によって支配されていたという。
また、ヘロドトスがしばしば「バクトリア人とサカ人」と並称させているところからみても、ペルシア人と先住のサカ人が共同して支配階級を構成していたものと思われる。
バクトリア・マルギアナ複合
http://ja.wikipedia.org/wiki/%E3%83%90%E3%82%AF%E3%83%88%E3%83%AA%E3%82%A2%E3%83%BB%E3%83%9E%E3%83%AB%E3%82%AE%E3%82%A2%E3%83%8A%E8%A4%87%E5%90%88
バクトリア・マルギアナ複合(-ふくごう;Bactria-Margiana Archeological Complex:略称BMAC)とは、
青銅器時代の紀元前2000年前後に、現在のトルクメニスタン、ウズベキスタン、タジキスタン、アフガニスタン北部のアムダリヤ(オクサス)川上流部などに栄えた
一連の先史文化を指す考古学用語である。
インダス文明とほぼ同時代に高度の都市文化を発展させたことから「第五の古代文明」という意味でオクサス文明とも呼ばれる。
発見は比較的新しく、研究途上にある。メソポタミアの文明やエラム文明、インダス文明など他の文化との関係、
特にアーリア人のインド・イランでの勃興に関連しても注目されている。
位置と時代
バクトリア(アフガニスタン北部)、マルギアナ(トルクメニスタン)は遺跡が集中する地域のギリシア語名である。
乾燥地帯であるが、川とオアシスを利用して古くから農業が行われた。
現在のメルヴを中心とし、アケメネス朝ペルシア以降栄えた。
BMACは、ソ連の考古学者ヴィクトル・サリアニディ(英版)が発掘調査に基づき1976年に命名した。
代表的な都市遺跡としてはナマズガ・デペ(英版)やアルティン・デペ(英語版)がある。
住民は灌漑により小麦・大麦などの栽培を行っていた。
都市や城塞の遺構のほかに、優れた金属器や、土器、宝石類、石の印章など様々な遺物が知られる。
印章に見られる図柄はイラン南東部から出土した陶器や銀器によく似ている。
マルグッシュ遺跡(英版)(ゴヌール・テペ)からはエラム文字と見られる銘文を彫った陶片が見つかっている。
これらの遺跡の上下限年代は、C14年代測定によって紀元前2200年から1500年頃と提示されている。
この発展と没落の過程はまだよくわかっていない。
BMACの遺物はこの地域だけでなくイラン東部、ペルシャ湾岸、バルチスタン、インダス川流域(ハラッパーなど)の広い範囲で見出されている。
中心地はアフガニスタン南部からバルチスタンにあったとする学者もいるが、
今のところ同地方から本格的な規模の同文化に属する遺跡は一切発見されていない。
http://ja.wikipedia.org/wiki/%E3%83%90%E3%82%AF%E3%83%88%E3%83%AA%E3%82%A2%E3%83%BB%E3%83%9E%E3%83%AB%E3%82%AE%E3%82%A2%E3%83%8A%E8%A4%87%E5%90%88
バクトリア・マルギアナ複合(-ふくごう;Bactria-Margiana Archeological Complex:略称BMAC)とは、
青銅器時代の紀元前2000年前後に、現在のトルクメニスタン、ウズベキスタン、タジキスタン、アフガニスタン北部のアムダリヤ(オクサス)川上流部などに栄えた
一連の先史文化を指す考古学用語である。
インダス文明とほぼ同時代に高度の都市文化を発展させたことから「第五の古代文明」という意味でオクサス文明とも呼ばれる。
発見は比較的新しく、研究途上にある。メソポタミアの文明やエラム文明、インダス文明など他の文化との関係、
特にアーリア人のインド・イランでの勃興に関連しても注目されている。
位置と時代
バクトリア(アフガニスタン北部)、マルギアナ(トルクメニスタン)は遺跡が集中する地域のギリシア語名である。
乾燥地帯であるが、川とオアシスを利用して古くから農業が行われた。
現在のメルヴを中心とし、アケメネス朝ペルシア以降栄えた。
BMACは、ソ連の考古学者ヴィクトル・サリアニディ(英版)が発掘調査に基づき1976年に命名した。
代表的な都市遺跡としてはナマズガ・デペ(英版)やアルティン・デペ(英語版)がある。
住民は灌漑により小麦・大麦などの栽培を行っていた。
都市や城塞の遺構のほかに、優れた金属器や、土器、宝石類、石の印章など様々な遺物が知られる。
印章に見られる図柄はイラン南東部から出土した陶器や銀器によく似ている。
マルグッシュ遺跡(英版)(ゴヌール・テペ)からはエラム文字と見られる銘文を彫った陶片が見つかっている。
これらの遺跡の上下限年代は、C14年代測定によって紀元前2200年から1500年頃と提示されている。
この発展と没落の過程はまだよくわかっていない。
BMACの遺物はこの地域だけでなくイラン東部、ペルシャ湾岸、バルチスタン、インダス川流域(ハラッパーなど)の広い範囲で見出されている。
中心地はアフガニスタン南部からバルチスタンにあったとする学者もいるが、
今のところ同地方から本格的な規模の同文化に属する遺跡は一切発見されていない。
>>945
他の文化との関係
イラク、イランやインダス地方とは盛んな交流が行われ交易圏を形成していたが、
別の独立した文明という説と、メソポタミアやエラム文明からの移住地として始まったという説がある。
東側では、土器などに関してガンダーラ墓葬文化(英語版)(GGC:スワート(Swat)文化ともいう)との深い関係が考えられている。
また同時期の北側では中央アジアの広い範囲に遊牧民のアンドロノヴォ文化が栄えており、これとの接触もあったようである。
サリアニディはBMAC文化の起源についてアナトリアなどに由来する説を称えているが、
イランのエラム文明に由来するとする説やイラクからインダス一帯の交易権圏の下で独自に発達したとする説、
他にほぼ同時期のタリム盆地の先史文化と結び付ける説もある。
アーリア人との関係
この時代は、アーリア人がインドやイランで勃興する直前の時期に当たり、BMACはこれとの関係でも注目されている。
アンドロノヴォ文化を原アーリア文化とする説があるが、この文化はインド・イランの考古学的文化と関連づけるのが難しい。
またアンドロノヴォ文化が原アーリア文化であれば、これがBMACを滅亡させたと想像されるが、
BMACは馬の牧畜と戦車を使用する文化により滅亡した形跡はあるものの、より北方に位置する地方の同文化の最も早い痕跡は紀元前1100年頃のものである。
またサリアニディ自身はBMAC=原アーリア説を主張し、大量の灰あるいはケシや麻黄が発見された宮殿の部屋をアーリア人の拝火儀式、ソーマ(ハオマ)儀式の証拠であるとするが、
BMACは農耕文化であって馬に関係した遺物は極めて乏しく、BMACを原アーリア人と関連づけるのは困難である。
またジェームズ・マロリー(英語版)はヴェーダにおける砦の記述と発掘された城塞とを結び付け、アンドロノヴォ文化がBMACと同化してアーリア文化になったと主張するが、
BMACとこれを滅亡させたと見られる文化は短期間かつ断絶的に入れ替わっている。
インド・イラン語派には印欧祖語やドラヴィダ語と異なる基層言語があるとの考えもあり、それがBMACの言語(単一ではない?)と考える人もいる。
現在ガンダーラ地方の近く(カシミール)に残っているブルシャスキー語も関係があるかもしれない。
他の文化との関係
イラク、イランやインダス地方とは盛んな交流が行われ交易圏を形成していたが、
別の独立した文明という説と、メソポタミアやエラム文明からの移住地として始まったという説がある。
東側では、土器などに関してガンダーラ墓葬文化(英語版)(GGC:スワート(Swat)文化ともいう)との深い関係が考えられている。
また同時期の北側では中央アジアの広い範囲に遊牧民のアンドロノヴォ文化が栄えており、これとの接触もあったようである。
サリアニディはBMAC文化の起源についてアナトリアなどに由来する説を称えているが、
イランのエラム文明に由来するとする説やイラクからインダス一帯の交易権圏の下で独自に発達したとする説、
他にほぼ同時期のタリム盆地の先史文化と結び付ける説もある。
アーリア人との関係
この時代は、アーリア人がインドやイランで勃興する直前の時期に当たり、BMACはこれとの関係でも注目されている。
アンドロノヴォ文化を原アーリア文化とする説があるが、この文化はインド・イランの考古学的文化と関連づけるのが難しい。
またアンドロノヴォ文化が原アーリア文化であれば、これがBMACを滅亡させたと想像されるが、
BMACは馬の牧畜と戦車を使用する文化により滅亡した形跡はあるものの、より北方に位置する地方の同文化の最も早い痕跡は紀元前1100年頃のものである。
またサリアニディ自身はBMAC=原アーリア説を主張し、大量の灰あるいはケシや麻黄が発見された宮殿の部屋をアーリア人の拝火儀式、ソーマ(ハオマ)儀式の証拠であるとするが、
BMACは農耕文化であって馬に関係した遺物は極めて乏しく、BMACを原アーリア人と関連づけるのは困難である。
またジェームズ・マロリー(英語版)はヴェーダにおける砦の記述と発掘された城塞とを結び付け、アンドロノヴォ文化がBMACと同化してアーリア文化になったと主張するが、
BMACとこれを滅亡させたと見られる文化は短期間かつ断絶的に入れ替わっている。
インド・イラン語派には印欧祖語やドラヴィダ語と異なる基層言語があるとの考えもあり、それがBMACの言語(単一ではない?)と考える人もいる。
現在ガンダーラ地方の近く(カシミール)に残っているブルシャスキー語も関係があるかもしれない。
>>945
http://upload.wikimedia.org/wikipedia/commons/b/b8/Indo-Iranian_origins.png
http://upload.wikimedia.org/wikipedia/commons/1/12/BMACAxeWithEagleHeadedDemonAndAnimals3rd-Early2ndMilleniumBCE.jpg
http://upload.wikimedia.org/wikipedia/commons/5/5a/BMACFemaleHead3rd-Early2ndMilleniumBCE.jpg
http://upload.wikimedia.org/wikipedia/commons/a/a4/BMACMonsterWithTrumpet3rd-Early2ndMilleniumBCE.jpg
http://upload.wikimedia.org/wikipedia/commons/b/b8/Indo-Iranian_origins.png
{kind=link}
http://upload.wikimedia.org/wikipedia/commons/1/12/BMACAxeWithEagleHeadedDemonAndAnimals3rd-Early2ndMilleniumBCE.jpg
{kind=link}
http://upload.wikimedia.org/wikipedia/commons/5/5a/BMACFemaleHead3rd-Early2ndMilleniumBCE.jpg
{kind=link}
http://upload.wikimedia.org/wikipedia/commons/a/a4/BMACMonsterWithTrumpet3rd-Early2ndMilleniumBCE.jpg
{kind=link}
マジャル語
http://ja.wikipedia.org/wiki/%E3%83%9E%E3%82%B8%E3%83%A3%E3%83%AB%E8%AA%9E
ウラル語族のフィン・ウゴル語派に分類され、フィンランド語やエストニア語と同系統の言語であるが、意思の疎通がまったくできないほどの大きな隔たりがある。
ヨーロッパで話される諸言語の多くが属するインド・ヨーロッパ語族とは系統が異なり(ただし歴史的経緯からスラヴ諸語やドイツ語の影響をある程度受けている)、
姓名や日付などの語順もインド・ヨーロッパ語族の言語とは異なる[1]。
ハンガリー
http://ja.wikipedia.org/wiki/%E3%83%8F%E3%83%B3%E3%82%AC%E3%83%AA%E3%83%BC
「ハンガリー」は俗説にあるような「フン族」が語源ではなく、「ウンガルン」(独: Ungarn)、「ウガリア」(希: Ουγγαρ?α)に見られるように元々は語頭の h がなかった。
語源として一般に認められているのは、7世紀のテュルク系の Onogur という語であり、十本の矢(十部族)を意味する。
これは初期のハンガリー人がマジャール7部族とハザール3部族の連合であったことに由来する。
http://ja.wikipedia.org/wiki/%E3%83%9E%E3%82%B8%E3%83%A3%E3%83%AB%E8%AA%9E
ウラル語族のフィン・ウゴル語派に分類され、フィンランド語やエストニア語と同系統の言語であるが、意思の疎通がまったくできないほどの大きな隔たりがある。
ヨーロッパで話される諸言語の多くが属するインド・ヨーロッパ語族とは系統が異なり(ただし歴史的経緯からスラヴ諸語やドイツ語の影響をある程度受けている)、
姓名や日付などの語順もインド・ヨーロッパ語族の言語とは異なる[1]。
ハンガリー
http://ja.wikipedia.org/wiki/%E3%83%8F%E3%83%B3%E3%82%AC%E3%83%AA%E3%83%BC
「ハンガリー」は俗説にあるような「フン族」が語源ではなく、「ウンガルン」(独: Ungarn)、「ウガリア」(希: Ουγγαρ?α)に見られるように元々は語頭の h がなかった。
語源として一般に認められているのは、7世紀のテュルク系の Onogur という語であり、十本の矢(十部族)を意味する。
これは初期のハンガリー人がマジャール7部族とハザール3部族の連合であったことに由来する。
>>948
ハンガリーの歴史
http://ja.wikipedia.org/wiki/%E3%83%8F%E3%83%B3%E3%82%AC%E3%83%AA%E3%83%BC%E3%81%AE%E6%AD%B4%E5%8F%B2
建国まで
この地域は、1世紀にはローマ帝国が属州パンノニアを置いた。4世紀にローマ帝国が衰退し、ゲルマン人の移動が始まると、この地にゴート族が到来した。
その後、ドナウ川中流の良質の平原に着目して、5世紀にフン族、6世紀にアヴァール人などテュルク系と見られる遊牧民の勢力がそれぞれ到来し定着した。
9世紀になるとウラル山脈の中南部周辺からウゴル系遊牧民であるマジャル人が黒海北岸〜ロシア南部のヴォルガ川南岸付近の広大な草原地帯に到達した。
その後、大首長(ジュラ)アールパードは共同統治者の名誉最高首長(ケンデ)クルサーンとともにバルカン半島を経由して、ハンガリー平野この地に移住してきた。
アールパードはパンノニア平原を根拠とし、アールパード家の祖となり、ドイツ南東部のバイエルンなど東・中部ヨーロッパ各地を騎馬で蹂躙し、同時期の海のヴァイキングとともに怖れられた。
しかし、955年にアールパードの孫タクショニュはレッヒフェルトの戦いでオットー1世に敗れると、
今までの部族の風習であるシャーマニズムによる自然崇拝を放棄し、カトリックによるキリスト教に改宗して、パンノニア平原に統一王国建設を開始した。
同時に現地のスラヴ族やラテン人など多くの民族と混血して、現地に同化した。
アヴァール
http://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%B4%E3%82%A1%E3%83%BC%E3%83%AB
概要
フンが姿を消してから約1世紀の後、フンと同じく現在のハンガリーの地を本拠に一大遊牧国家を築いたのがアヴァールである。
フンほどの強大さはなく、またアッティラほど傑出した指導者がいたわけでもなく、さらに周辺民族による記録が少なかったためにアヴァールの歴史はよく知られていない。
しかし、アヴァールは東ローマ帝国およびフランク王国と接触し、スラヴ諸民族の形成に大きな影響を与えた。[1]
言語系統[編集]
アヴァールの言語は「テュルク系」説、「柔然と同族なのでモンゴル系」説、の2つがあるがどちらなのかは不明である。
ハンガリーの歴史
http://ja.wikipedia.org/wiki/%E3%83%8F%E3%83%B3%E3%82%AC%E3%83%AA%E3%83%BC%E3%81%AE%E6%AD%B4%E5%8F%B2
建国まで
この地域は、1世紀にはローマ帝国が属州パンノニアを置いた。4世紀にローマ帝国が衰退し、ゲルマン人の移動が始まると、この地にゴート族が到来した。
その後、ドナウ川中流の良質の平原に着目して、5世紀にフン族、6世紀にアヴァール人などテュルク系と見られる遊牧民の勢力がそれぞれ到来し定着した。
9世紀になるとウラル山脈の中南部周辺からウゴル系遊牧民であるマジャル人が黒海北岸〜ロシア南部のヴォルガ川南岸付近の広大な草原地帯に到達した。
その後、大首長(ジュラ)アールパードは共同統治者の名誉最高首長(ケンデ)クルサーンとともにバルカン半島を経由して、ハンガリー平野この地に移住してきた。
アールパードはパンノニア平原を根拠とし、アールパード家の祖となり、ドイツ南東部のバイエルンなど東・中部ヨーロッパ各地を騎馬で蹂躙し、同時期の海のヴァイキングとともに怖れられた。
しかし、955年にアールパードの孫タクショニュはレッヒフェルトの戦いでオットー1世に敗れると、
今までの部族の風習であるシャーマニズムによる自然崇拝を放棄し、カトリックによるキリスト教に改宗して、パンノニア平原に統一王国建設を開始した。
同時に現地のスラヴ族やラテン人など多くの民族と混血して、現地に同化した。
アヴァール
http://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%B4%E3%82%A1%E3%83%BC%E3%83%AB
概要
フンが姿を消してから約1世紀の後、フンと同じく現在のハンガリーの地を本拠に一大遊牧国家を築いたのがアヴァールである。
フンほどの強大さはなく、またアッティラほど傑出した指導者がいたわけでもなく、さらに周辺民族による記録が少なかったためにアヴァールの歴史はよく知られていない。
しかし、アヴァールは東ローマ帝国およびフランク王国と接触し、スラヴ諸民族の形成に大きな影響を与えた。[1]
言語系統[編集]
アヴァールの言語は「テュルク系」説、「柔然と同族なのでモンゴル系」説、の2つがあるがどちらなのかは不明である。
>>949
ハンガリー王国[編集]
聖イシュトヴァーンの王冠と宝器1000年にタクショニュの孫であるアールパード家のイシュトヴァーン1世が本格的にキリスト教に改宗すると、
ローマ教皇からハンガリー国王として聖別をうけ戴冠し、ヨーロッパ世界の一員となった。
それからのハンガリー王国は北部のスロヴァキア(モラヴィア)、南部のクロアチアのスラヴ人を支配下に入れ、さらにルーマニアトランシルヴァニアにも勢力を伸ばした。
この頃がハンガリーの絶頂期であり、中欧の強国として君臨していた。
この時代の領域は聖イシュトヴァーンの王冠の地と呼ばれ、以後ハンガリーの歴史観において重要な位置を占めた。
このためハンガリー王となるものは聖イシュトヴァーンの王冠を頂く者であるという概念が生まれた。
1240年にはモンゴル帝国のバトゥによる侵略を受け、甚大な被害をうけた(モンゴルのポーランド侵攻)。
この経験を経たことでハンガリー国王は防衛体制を整える必要に迫られ、貴族層に土地を与えて彼らの主導で堅固な城塞を築かせていった。
同じく防衛上の観点からも城壁を持つ都市の発展が求められ、従来までの都市のほか、新たにドイツ人の入植を契機とした都市も形成・発展した。
その例として、シビウ、ブラショフ、ビストリツァ、コシツェなどが挙げられる。
ハンガリー王国[編集]
聖イシュトヴァーンの王冠と宝器1000年にタクショニュの孫であるアールパード家のイシュトヴァーン1世が本格的にキリスト教に改宗すると、
ローマ教皇からハンガリー国王として聖別をうけ戴冠し、ヨーロッパ世界の一員となった。
それからのハンガリー王国は北部のスロヴァキア(モラヴィア)、南部のクロアチアのスラヴ人を支配下に入れ、さらにルーマニアトランシルヴァニアにも勢力を伸ばした。
この頃がハンガリーの絶頂期であり、中欧の強国として君臨していた。
この時代の領域は聖イシュトヴァーンの王冠の地と呼ばれ、以後ハンガリーの歴史観において重要な位置を占めた。
このためハンガリー王となるものは聖イシュトヴァーンの王冠を頂く者であるという概念が生まれた。
1240年にはモンゴル帝国のバトゥによる侵略を受け、甚大な被害をうけた(モンゴルのポーランド侵攻)。
この経験を経たことでハンガリー国王は防衛体制を整える必要に迫られ、貴族層に土地を与えて彼らの主導で堅固な城塞を築かせていった。
同じく防衛上の観点からも城壁を持つ都市の発展が求められ、従来までの都市のほか、新たにドイツ人の入植を契機とした都市も形成・発展した。
その例として、シビウ、ブラショフ、ビストリツァ、コシツェなどが挙げられる。
>>949
東ローマ帝国との同盟[編集]
アヴァールが歴史上に現れるのは558年のことで、時に東ローマ帝国ではユスティニアヌス1世(在位:518年 - 565年)の治世であった。
アヴァールは突厥に追われて北カフカスに姿を現し、アラン人の仲介で東ローマ帝国と同盟関係を結んだ。
567年、アヴァールはゲルマン系のランゴバルド人と組み、ダキアとトランシルヴァニア、東パンノニアに割拠していたゲルマン系のゲピド族を滅ぼし、
その地を奪った(アヴァール可汗国(英語版)[7]の建国)。
翌年(568年)、ランゴバルドがイタリア半島に向かいランゴバルド王国を建国すると、アヴァールはそれに代わってハンガリー平原全域を支配した。
ここにおいてアヴァールの勢力範囲は、ティサ川流域を中心にボヘミアからドナウ川流域を経て南ロシアにおよぶ広大なものとなった。
この年、突厥可汗国の室点蜜(Stembis)の使者がコンスタンティノープルに現れ、東ローマ帝国と対ペルシア同盟を組み友好関係を結んだ。
東ローマ帝国ではユスティニアヌス1世が死去し、ユスティヌス2世(在位:565年 - 578年)が即位していた。
ユスティヌス2世はアヴァールに対して強硬姿勢を執り、アヴァールの使節に対して貢納の支払いを拒否したが、
アヴァールの指導者バヤン・カガン(英語版)の怒りを買い、バルカン半島の要衝であるサヴァ川沿いの要塞シルミウムを陥落寸前までに追い込まれた。
これによって、ユスティヌス2世は574年にアヴァールへの貢納を再開することとなる。
東ローマ帝国と突厥可汗国は568年以来、使節を往来させていたが、ふたたび東ローマがアヴァールと同盟を組んだことで両者の関係が一気に崩れ、
576年に突厥は東ローマの使節を非難するとともに(突厥はかつて自分たちが打ち破ったアヴァール人と同盟を結んだことに不信感を抱いた)クリミア半島の東ローマ領を征服した。
東ローマ帝国との同盟[編集]
アヴァールが歴史上に現れるのは558年のことで、時に東ローマ帝国ではユスティニアヌス1世(在位:518年 - 565年)の治世であった。
アヴァールは突厥に追われて北カフカスに姿を現し、アラン人の仲介で東ローマ帝国と同盟関係を結んだ。
567年、アヴァールはゲルマン系のランゴバルド人と組み、ダキアとトランシルヴァニア、東パンノニアに割拠していたゲルマン系のゲピド族を滅ぼし、
その地を奪った(アヴァール可汗国(英語版)[7]の建国)。
翌年(568年)、ランゴバルドがイタリア半島に向かいランゴバルド王国を建国すると、アヴァールはそれに代わってハンガリー平原全域を支配した。
ここにおいてアヴァールの勢力範囲は、ティサ川流域を中心にボヘミアからドナウ川流域を経て南ロシアにおよぶ広大なものとなった。
この年、突厥可汗国の室点蜜(Stembis)の使者がコンスタンティノープルに現れ、東ローマ帝国と対ペルシア同盟を組み友好関係を結んだ。
東ローマ帝国ではユスティニアヌス1世が死去し、ユスティヌス2世(在位:565年 - 578年)が即位していた。
ユスティヌス2世はアヴァールに対して強硬姿勢を執り、アヴァールの使節に対して貢納の支払いを拒否したが、
アヴァールの指導者バヤン・カガン(英語版)の怒りを買い、バルカン半島の要衝であるサヴァ川沿いの要塞シルミウムを陥落寸前までに追い込まれた。
これによって、ユスティヌス2世は574年にアヴァールへの貢納を再開することとなる。
東ローマ帝国と突厥可汗国は568年以来、使節を往来させていたが、ふたたび東ローマがアヴァールと同盟を組んだことで両者の関係が一気に崩れ、
576年に突厥は東ローマの使節を非難するとともに(突厥はかつて自分たちが打ち破ったアヴァール人と同盟を結んだことに不信感を抱いた)クリミア半島の東ローマ領を征服した。
>>949
アヴァール対スラヴ[編集]
623年頃、最初のスラヴ国家であるサモ王国(英語版)(623年-658年)が旧チェコスロヴァキアの地に形成され、その地のスラヴ人がアヴァールの支配を脱した。
サモ王国は7世紀後半にアヴァールによって滅ぼされるが、すでにアヴァールの方も衰退期に入っており、全体としてはスラヴ人が独立性を強めていった。
791年、フランクのカール大帝がアヴァールに遠征し、804年までにドナウ川中流域を征服。
一方で南のブルガールもアヴァールを追ってパンノニアまで進出したため、アヴァールはフランク,ブルガール,スラヴの3者によって分割され滅亡した。
アヴァールの国家組織[編集]
アヴァール可汗国は強力な軍事力と発達した政治機構を持つ遊牧国家であり、支配者は遊牧国家の君主号であるカガン(khagan:可汗)を称した。
カガンを中心として「イウグル」と「トゥドゥン」と呼ばれる二人の高官が補佐する体制であったとされる。
またパンノニアで発見されたアヴァールが残したと考えられる鐙・火打ち金などの出土品は東アジアや北アジアに起源があり、
アヴァールが鐙を西欧に伝えたことで西欧の戦闘法に大きな影響を与えた。
一方で、アヴァール人の進出によってカルパチア盆地やドナウ川上流域に残っていたテウルニア、ウィルーヌムといった司教区は消滅した。
アヴァール対スラヴ[編集]
623年頃、最初のスラヴ国家であるサモ王国(英語版)(623年-658年)が旧チェコスロヴァキアの地に形成され、その地のスラヴ人がアヴァールの支配を脱した。
サモ王国は7世紀後半にアヴァールによって滅ぼされるが、すでにアヴァールの方も衰退期に入っており、全体としてはスラヴ人が独立性を強めていった。
791年、フランクのカール大帝がアヴァールに遠征し、804年までにドナウ川中流域を征服。
一方で南のブルガールもアヴァールを追ってパンノニアまで進出したため、アヴァールはフランク,ブルガール,スラヴの3者によって分割され滅亡した。
アヴァールの国家組織[編集]
アヴァール可汗国は強力な軍事力と発達した政治機構を持つ遊牧国家であり、支配者は遊牧国家の君主号であるカガン(khagan:可汗)を称した。
カガンを中心として「イウグル」と「トゥドゥン」と呼ばれる二人の高官が補佐する体制であったとされる。
またパンノニアで発見されたアヴァールが残したと考えられる鐙・火打ち金などの出土品は東アジアや北アジアに起源があり、
アヴァールが鐙を西欧に伝えたことで西欧の戦闘法に大きな影響を与えた。
一方で、アヴァール人の進出によってカルパチア盆地やドナウ川上流域に残っていたテウルニア、ウィルーヌムといった司教区は消滅した。
>>745
さて、フンガリア
ウクライナ、クロアチア、チェコ の真ん中にいるかな。
トルコは クロアチア、ブルガリア辺りに枝が見える。
それにしても、シリアとトルコの間が遠すぎる。
レヴァントの回廊を通らないと繋がらない、、、
さて、フンガリア
ウクライナ、クロアチア、チェコ の真ん中にいるかな。
トルコは クロアチア、ブルガリア辺りに枝が見える。
それにしても、シリアとトルコの間が遠すぎる。
レヴァントの回廊を通らないと繋がらない、、、
>>761
ところでこの年代はどこから判明したの?
まだ見逃してる遺跡があるとか?
> とにかくR1bが欧州に侵入したのは4500〜5000年前だからね。
> それまでに狩猟採集民と農耕民の最低ふたつの人類の流れがあるわけだ。
ところでこの年代はどこから判明したの?
まだ見逃してる遺跡があるとか?
> とにかくR1bが欧州に侵入したのは4500〜5000年前だからね。
> それまでに狩猟採集民と農耕民の最低ふたつの人類の流れがあるわけだ。
あっちに長く住んで白人を見慣れれば美男美女ばかりじゃないことに気づくよ
956 :世界@名無史さん:2014/02/12(水) 19:38:46.76 0
>>956
書いてあるのは見つからなかったけど
該当物はありました
http://r1b.org/?page_id=112
R1b and the Bell Beaker Phenomenon
Ancient DNA analysis of two male skeletons from the Late Neolithic Bell Beaker site of Kromsdorf, Germany showed they belonged to Y-DNA haplogroup R1b.[1]
More specifically, one skeleton belonged to R1b (M343) with the testing of R1b1a2 (marker M269) having failed and the other skeleton belonged to R1b1a2.
Both were ancestral for SNP U106. No other downstream markers were tested.
The find is important because it links the widespread Bell Beaker Phenomenon (hereafter BB) with the most frequent Y-DNA haplogroup in modern Western European males.
It is also important as R1b has not appeared in any Neolithic or pre-Neolithic ancient DNA to date.
Resources
1. Lee, E. et al. (2012), Emerging genetic patterns of the European neolithic: Perspectives from a late neolithic bell beaker burial site in Germany,
American Journal of Physical Anthropology, online 3 May 2012 ahead of print.
2. Muller J, Van Willigen S, New radiocarbon evidence for European Bell Beakers and the consequences for the diffusion of the Bell Beaker Phenomenon, in Franco Nicolis (ed.), Bell Beakers today:
Pottery, people, culture, symbols in prehistoric Europe (2001), pp. 59-75.
3. Vander Linden M, Demography and mobility in North-Western Europe during the third millennium cal.
BC, Prescott C. & Glorstad H. (reds.), Becoming European. The transformation of third millennium Europe and the trajectory into the millennium BC (2012). Oxbow Books, Oxford, p. 22.
書いてあるのは見つからなかったけど
該当物はありました
http://r1b.org/?page_id=112
R1b and the Bell Beaker Phenomenon
Ancient DNA analysis of two male skeletons from the Late Neolithic Bell Beaker site of Kromsdorf, Germany showed they belonged to Y-DNA haplogroup R1b.[1]
More specifically, one skeleton belonged to R1b (M343) with the testing of R1b1a2 (marker M269) having failed and the other skeleton belonged to R1b1a2.
Both were ancestral for SNP U106. No other downstream markers were tested.
The find is important because it links the widespread Bell Beaker Phenomenon (hereafter BB) with the most frequent Y-DNA haplogroup in modern Western European males.
It is also important as R1b has not appeared in any Neolithic or pre-Neolithic ancient DNA to date.
Resources
1. Lee, E. et al. (2012), Emerging genetic patterns of the European neolithic: Perspectives from a late neolithic bell beaker burial site in Germany,
American Journal of Physical Anthropology, online 3 May 2012 ahead of print.
2. Muller J, Van Willigen S, New radiocarbon evidence for European Bell Beakers and the consequences for the diffusion of the Bell Beaker Phenomenon, in Franco Nicolis (ed.), Bell Beakers today:
Pottery, people, culture, symbols in prehistoric Europe (2001), pp. 59-75.
3. Vander Linden M, Demography and mobility in North-Western Europe during the third millennium cal.
BC, Prescott C. & Glorstad H. (reds.), Becoming European. The transformation of third millennium Europe and the trajectory into the millennium BC (2012). Oxbow Books, Oxford, p. 22.
958 :世界@名無史さん:2014/02/12(水) 21:18:17.91 0
>>957
ありがとう。
このくだりだね。
It is also important as R1b has not appeared in any Neolithic or pre-Neolithic ancient DNA to date.
ここ1〜2年の間に古代欧州人の変遷というか血統の入れ替わりに関する記事・論文が増えたね。
最後に登場するのがR1bということなんだが。
ありがとう。
このくだりだね。
It is also important as R1b has not appeared in any Neolithic or pre-Neolithic ancient DNA to date.
ここ1〜2年の間に古代欧州人の変遷というか血統の入れ替わりに関する記事・論文が増えたね。
最後に登場するのがR1bということなんだが。
ウーニェチツェ文化
http://ja.wikipedia.org/wiki/%E3%82%A6%E3%83%BC%E3%83%8B%E3%82%A7%E3%83%81%E3%83%84%E3%82%A7%E6%96%87%E5%8C%96
ウーニェチツェ文化(英語:Un?tice culture)は紀元前2300年から紀元前1600年にかけての中央ヨーロッパの青銅器時代の考古文化。
名称はプラハの北東郊外にこの文化の遺跡があるウーニェチツェ地区から採られた。中核部はチェコ、ドイツ中部と南部、ポーランド西部と中部に広がっている。
ウーニェチツェ文化は、先行する縄目文土器文化にとって代わって広がった。その後西部と東部では異なる発展段階を辿ることになる:
http://upload.wikimedia.org/wikipedia/commons/f/f9/Central_Europe_Reinecke_BA1.png
この文化は、パウル・ライネッケ(en:Paul Reinecke)の年代区分では青銅器時代A1とA2に相当する:
A1:2300-1950 BC:三角ダガー、平斧、石製のリストガード(en:stone wrist-guards)、フリント製の鏃
A2:1950-1700 BC:金属製の柄の付いたダガー、フランジ付き斧(フランジ斧)、ハルバード、球形の頭部に穿孔のある針類、堅固なブレスレット
これらの区分年代は主にジンゲン集団墓地(放射性炭素年代測定)、およびレウビンゲン墓地とヘルムスドルフ墓地(年輪年代測定)での調査による。
マリヤ・ギンブタスによると、この文化の墓地遺跡にはバルト海産の琥珀が高い割合で埋められているという。
有名なネブラ・ディスクは、ウーニェチツェ文化のものとされる。この文化に特徴的な銅製のダガーが、この円盤とともに複数発見されたことによる。
http://ja.wikipedia.org/wiki/%E3%82%A6%E3%83%BC%E3%83%8B%E3%82%A7%E3%83%81%E3%83%84%E3%82%A7%E6%96%87%E5%8C%96
ウーニェチツェ文化(英語:Un?tice culture)は紀元前2300年から紀元前1600年にかけての中央ヨーロッパの青銅器時代の考古文化。
名称はプラハの北東郊外にこの文化の遺跡があるウーニェチツェ地区から採られた。中核部はチェコ、ドイツ中部と南部、ポーランド西部と中部に広がっている。
ウーニェチツェ文化は、先行する縄目文土器文化にとって代わって広がった。その後西部と東部では異なる発展段階を辿ることになる:
http://upload.wikimedia.org/wikipedia/commons/f/f9/Central_Europe_Reinecke_BA1.png
{kind=link}
この文化は、パウル・ライネッケ(en:Paul Reinecke)の年代区分では青銅器時代A1とA2に相当する:
A1:2300-1950 BC:三角ダガー、平斧、石製のリストガード(en:stone wrist-guards)、フリント製の鏃
A2:1950-1700 BC:金属製の柄の付いたダガー、フランジ付き斧(フランジ斧)、ハルバード、球形の頭部に穿孔のある針類、堅固なブレスレット
これらの区分年代は主にジンゲン集団墓地(放射性炭素年代測定)、およびレウビンゲン墓地とヘルムスドルフ墓地(年輪年代測定)での調査による。
マリヤ・ギンブタスによると、この文化の墓地遺跡にはバルト海産の琥珀が高い割合で埋められているという。
有名なネブラ・ディスクは、ウーニェチツェ文化のものとされる。この文化に特徴的な銅製のダガーが、この円盤とともに複数発見されたことによる。
>>959
死亡推定ワラタww
交易[編集]
ウーニェチツェ文化の交易相手の文化のなかには、ストーンヘンジで有名なイギリスの先史時代社会(Wessex culture)も含まれていた。
ウーニェチツェ文化の鍛冶屋たちは純粋な銅を使っていた。ヒ素・アンチモン・錫の銅合金によって青銅を作るのはこれよりもあとの時代からである。
例外もあり、たとえば先に挙げたジンゲンの集団墓地では、錫を9%混ぜた銅合金のダガーが何本か見つかっている。
このダガーはおそらくブルターニュ半島で作られたものと見られる。なお、ブルターニュ半島でもその地方の富裕層の墓が数か所見つかっている。
アイルランド産の錫は広く取引されていた。ドイツ・ヘッセン州のブツバッハ(Butzbach)では、リューニャラ(lunula)と呼ばれる三日月形ネックレスが見つかっているが、これは金製でアイルランド風の意匠となっている。
琥珀も取引されていた。世界的な大産地であるバルト海南岸産の琥珀(バルチック・アンバー)のほか、他所の小規模な産地で採られた琥珀も取引されていたようである。
このように周辺各地との人的交流は盛んで、最近ではスウェーデンのスコーネ地方から来たと思われる男性の遺骨がポーランドのヴロツワフ市で見つかっている。
この「スウェーデン人」男性は、滞在していたこの土地で地元の「ポーランド人」女性2人に同時に手を出したことが村人たちに発覚して捕えられ、女性たちと共に処刑されたものと推定される。[1]
死亡推定ワラタww
交易[編集]
ウーニェチツェ文化の交易相手の文化のなかには、ストーンヘンジで有名なイギリスの先史時代社会(Wessex culture)も含まれていた。
ウーニェチツェ文化の鍛冶屋たちは純粋な銅を使っていた。ヒ素・アンチモン・錫の銅合金によって青銅を作るのはこれよりもあとの時代からである。
例外もあり、たとえば先に挙げたジンゲンの集団墓地では、錫を9%混ぜた銅合金のダガーが何本か見つかっている。
このダガーはおそらくブルターニュ半島で作られたものと見られる。なお、ブルターニュ半島でもその地方の富裕層の墓が数か所見つかっている。
アイルランド産の錫は広く取引されていた。ドイツ・ヘッセン州のブツバッハ(Butzbach)では、リューニャラ(lunula)と呼ばれる三日月形ネックレスが見つかっているが、これは金製でアイルランド風の意匠となっている。
琥珀も取引されていた。世界的な大産地であるバルト海南岸産の琥珀(バルチック・アンバー)のほか、他所の小規模な産地で採られた琥珀も取引されていたようである。
このように周辺各地との人的交流は盛んで、最近ではスウェーデンのスコーネ地方から来たと思われる男性の遺骨がポーランドのヴロツワフ市で見つかっている。
この「スウェーデン人」男性は、滞在していたこの土地で地元の「ポーランド人」女性2人に同時に手を出したことが村人たちに発覚して捕えられ、女性たちと共に処刑されたものと推定される。[1]
>>958
年代はその続きにあるけど
元は確認してなす
>>957
Based on the radiocarbon (14C) dating of short lived material, the current prevailing view is that BB originated in Iberia (2900 BC cal.),
with an almost concurrent appearance in southern France and northern Italy.[2]
The spread of BB into Northern and Central Europe seems to have occurred somewhat later (~2500 BC).
Vander Linden (2012) questioned the use of 14C dating to find the origins of BB, mainly on the grounds that most dates fall within a very narrow time-frame.[3]
He reinforced instead the Dutch Model, which based on typology and burial data, sees BB as an evolution of the Single Grace Culture (Corded Ware) in the lower Rhine.
Limited ancient DNA has failed to provide a male genetic link however as Corded Ware skeletons
have been found to belong instead to haplogroup R1a1, haplogroup G and possibly haplogroup I. [4] [5]
Based on the homogeneity of STR variance of the three major subclades of P312 (U152, L21 and DF27), and similar modal values for U106 (65 of 67),[6]
both P312 and U106 seem to have had a great period of geographic expansion in a relatively short period of time.
A second, albeit less frequent, brother clade to L11 is defined by Y chromosome position 3263086 G>A (GRCh37/hg19 Assembly) (unpublished data).
The distribution of this group is primarily restricted to Northern Italy and France[6] and is easily spotted in academic studies by way of its off-modal value of DYS426=13.
While BB migrations seem to have impacted some areas such as Bavaria very heavily[7],
other areas such as Northern Iberia and Bohemia show very little variability from the preceding non-BB populations [8].
Understanding the dynamics of Bell Beaker population movements and how they shaped the distribution of R1b
and its phylogeny should warrant Y-DNA testing of ancient skeletal remains at the subclade level.[9]
年代はその続きにあるけど
元は確認してなす
>>957
Based on the radiocarbon (14C) dating of short lived material, the current prevailing view is that BB originated in Iberia (2900 BC cal.),
with an almost concurrent appearance in southern France and northern Italy.[2]
The spread of BB into Northern and Central Europe seems to have occurred somewhat later (~2500 BC).
Vander Linden (2012) questioned the use of 14C dating to find the origins of BB, mainly on the grounds that most dates fall within a very narrow time-frame.[3]
He reinforced instead the Dutch Model, which based on typology and burial data, sees BB as an evolution of the Single Grace Culture (Corded Ware) in the lower Rhine.
Limited ancient DNA has failed to provide a male genetic link however as Corded Ware skeletons
have been found to belong instead to haplogroup R1a1, haplogroup G and possibly haplogroup I. [4] [5]
Based on the homogeneity of STR variance of the three major subclades of P312 (U152, L21 and DF27), and similar modal values for U106 (65 of 67),[6]
both P312 and U106 seem to have had a great period of geographic expansion in a relatively short period of time.
A second, albeit less frequent, brother clade to L11 is defined by Y chromosome position 3263086 G>A (GRCh37/hg19 Assembly) (unpublished data).
The distribution of this group is primarily restricted to Northern Italy and France[6] and is easily spotted in academic studies by way of its off-modal value of DYS426=13.
While BB migrations seem to have impacted some areas such as Bavaria very heavily[7],
other areas such as Northern Iberia and Bohemia show very little variability from the preceding non-BB populations [8].
Understanding the dynamics of Bell Beaker population movements and how they shaped the distribution of R1b
and its phylogeny should warrant Y-DNA testing of ancient skeletal remains at the subclade level.[9]
962 :世界@名無史さん:2014/02/12(水) 22:32:30.24 0
英語で書き込んでくれている人、ありがとう。
今度の土日にじっくり読んで考えるわ。
フィナンシャルタイムズよりはるかに面白い。
今度の土日にじっくり読んで考えるわ。
フィナンシャルタイムズよりはるかに面白い。
>>957
http://dienekes.blogspot.jp/2012/05/bell-beakers-from-germany-y-haplogroup.html
http://onlinelibrary.wiley.com/doi/10.1002/ajpa.22074/abstract
American Journal of Physical Anthropology DOI: 10.1002/ajpa.22074
Emerging genetic patterns of the european neolithic: Perspectives from a late neolithic bell beaker burial site in Germany†
The transition from hunting and gathering to agriculture in Europe is associated with demographic changes that may have shifted the human gene pool of the region as a result of an influx of Neolithic farmers from the Near East.
However, the genetic composition of populations after the earliest Neolithic,
when a diverse mosaic of societies that had been fully engaged in agriculture for some time appeared in central Europe, is poorly known.
At this period during the Late Neolithic (ca. 2,800?2,000 BC), regionally distinctive burial patterns associated with two different cultural groups emerge, Bell Beaker and Corded Ware,
and may reflect differences in how these societies were organized. Ancient DNA analyses of human remains from the Late Neolithic Bell Beaker site of Kromsdorf, Germany
showed distinct mitochondrial haplotypes for six individuals, which were classified under
the haplogroups I1, K1, T1, U2, U5, and W5, and two males were identified as belonging to the Y haplogroup R1b.
In contrast to other Late Neolithic societies in Europe emphasizing maintenance of biological relatedness in mortuary contexts,
the diversity of maternal haplotypes evident at Kromsdorf suggests that burial practices of Bell Beaker communities operated outside of social norms based on shared maternal lineages.
Furthermore, our data, along with those from previous studies, indicate that modern U5-lineages may have received little, if any, contribution from the Mesolithic or Neolithic mitochondrial gene pool.
http://dienekes.blogspot.jp/2012/05/bell-beakers-from-germany-y-haplogroup.html
http://onlinelibrary.wiley.com/doi/10.1002/ajpa.22074/abstract
American Journal of Physical Anthropology DOI: 10.1002/ajpa.22074
Emerging genetic patterns of the european neolithic: Perspectives from a late neolithic bell beaker burial site in Germany†
The transition from hunting and gathering to agriculture in Europe is associated with demographic changes that may have shifted the human gene pool of the region as a result of an influx of Neolithic farmers from the Near East.
However, the genetic composition of populations after the earliest Neolithic,
when a diverse mosaic of societies that had been fully engaged in agriculture for some time appeared in central Europe, is poorly known.
At this period during the Late Neolithic (ca. 2,800?2,000 BC), regionally distinctive burial patterns associated with two different cultural groups emerge, Bell Beaker and Corded Ware,
and may reflect differences in how these societies were organized. Ancient DNA analyses of human remains from the Late Neolithic Bell Beaker site of Kromsdorf, Germany
showed distinct mitochondrial haplotypes for six individuals, which were classified under
the haplogroups I1, K1, T1, U2, U5, and W5, and two males were identified as belonging to the Y haplogroup R1b.
In contrast to other Late Neolithic societies in Europe emphasizing maintenance of biological relatedness in mortuary contexts,
the diversity of maternal haplotypes evident at Kromsdorf suggests that burial practices of Bell Beaker communities operated outside of social norms based on shared maternal lineages.
Furthermore, our data, along with those from previous studies, indicate that modern U5-lineages may have received little, if any, contribution from the Mesolithic or Neolithic mitochondrial gene pool.
>>959
Nebra sky disk
http://en.wikipedia.org/wiki/Nebra_sky_disk
Origin of the metals[edit]
According to an initial analysis of trace elements by x-ray fluorescence by E. Pernicka, then at the University of Freiberg,
the copper originated at Bischofshofen in Austria,
while the gold was thought to be from the Carpathian Mountains.[3]
A more recent analysis found that the gold used in the first phase was from the river Carnon in Cornwall.[4]
The tin content of the bronze was also from Cornwall.[5]
Nebra sky disk
http://en.wikipedia.org/wiki/Nebra_sky_disk
Origin of the metals[edit]
According to an initial analysis of trace elements by x-ray fluorescence by E. Pernicka, then at the University of Freiberg,
the copper originated at Bischofshofen in Austria,
while the gold was thought to be from the Carpathian Mountains.[3]
A more recent analysis found that the gold used in the first phase was from the river Carnon in Cornwall.[4]
The tin content of the bronze was also from Cornwall.[5]
lie Teviec
http://en.wikipedia.org/wiki/Prehistory_of_Brittany
Mesolithic[edit]The best-known mesolithic sites from Brittany are the cemeteries on the islands of Hoedic (10 graves) and Teviec (9 graves) in Morbihan.
The collective graves are placed in shell middens without any particular order. Some graves show evidence of postmortal manipulations of the bones.
There are single burials and empty graves (cenotaphs) as well. The graves are covered with stones, a hearth or antlers forming a sort of dome.
Rich funeral gifts, flint tools, engraved bones, shell ornaments and ochre demonstrate the affluence of these hunter-gatherers, or rather fisher-gatherers. Certain shells are sex-specific.
In Teviec there are stone cist graves. The bones of an infant have been postmortally ornamented with striations.
The corresponding settlements consist of shell middens.
A radiocarbon date of 4625 (uncal.) for Hoedic places it in the 6th Millennium BC cal, rather late in the Mesolithic sequence,
and indeed there are some indications of contact with agricultural societies to the East.
Their economy was based on marine resources. Recently, a number of accelerator dates have been published for Hoedic.
In Beg an Dorchenn in Plomeur (Finistere), domestic dog and cattle were already present, in Dissignac, microliths were associated with pollen evidence for clearances.
Some scholars speculate that megalithic graves might go back to the Mesolithic, but this contention is difficult to prove,
as most structures have been reused. Large numbers of microliths have been found under the chambered tomb of Dissignac fr:Tumulus de Dissignac.
http://en.wikipedia.org/wiki/Prehistory_of_Brittany
Mesolithic[edit]The best-known mesolithic sites from Brittany are the cemeteries on the islands of Hoedic (10 graves) and Teviec (9 graves) in Morbihan.
The collective graves are placed in shell middens without any particular order. Some graves show evidence of postmortal manipulations of the bones.
There are single burials and empty graves (cenotaphs) as well. The graves are covered with stones, a hearth or antlers forming a sort of dome.
Rich funeral gifts, flint tools, engraved bones, shell ornaments and ochre demonstrate the affluence of these hunter-gatherers, or rather fisher-gatherers. Certain shells are sex-specific.
In Teviec there are stone cist graves. The bones of an infant have been postmortally ornamented with striations.
The corresponding settlements consist of shell middens.
A radiocarbon date of 4625 (uncal.) for Hoedic places it in the 6th Millennium BC cal, rather late in the Mesolithic sequence,
and indeed there are some indications of contact with agricultural societies to the East.
Their economy was based on marine resources. Recently, a number of accelerator dates have been published for Hoedic.
In Beg an Dorchenn in Plomeur (Finistere), domestic dog and cattle were already present, in Dissignac, microliths were associated with pollen evidence for clearances.
Some scholars speculate that megalithic graves might go back to the Mesolithic, but this contention is difficult to prove,
as most structures have been reused. Large numbers of microliths have been found under the chambered tomb of Dissignac fr:Tumulus de Dissignac.
>>928
ヌーリスターン語派
http://ja.wikipedia.org/wiki/%E3%83%8C%E3%83%BC%E3%83%AA%E3%82%B9%E3%82%BF%E3%83%BC%E3%83%B3%E8%AA%9E%E6%B4%BE
ヌーリスターン語派(Nuristan, ヌーリスタン語派)、またはカーフィル語派(Kafir)とは、
パキスタンからアフガニスタンにかけて分布する言語のグループで、インド・ヨーロッパ語族のインド・イラン語派に属する。
比較言語学的な位置づけについては、近年ではインド・イラン語派内の独立した一グループとする見解が主流であるが、
一方で支持は少ないながらも、インド語派に含める説や、イラン語派に属するがインド語派に属するダルド語派から多大な影響を受けて変容したとする説もある。
いずれにせよ、ヌーリスターン語派の諸民族が現住地に落ち着いたのは相当の過去のことであり、またインド語派とは異なりパンジャーブ地方へは入らなかったという点に争いはない。
ヌーリスターンは1890年代までペルシア語で「不信仰者の地」を意味するカフィリスタン(英語版)と呼ばれていた。
これは当地の住民が独自の多神教(アニミズム)を信仰していたためで、言語もこれに倣ってカーフィル語派とされていた。
しかしアブドゥッラフマーン・ハーンに征服されて強制的に改宗させられ、住民の一部がイスラームを受容すると、
異教の民がイスラームの光に照らされたとしてヌーリスターン(光の地)と呼ばれるようになり、
カーフィル語派の名も差別的だとしてヌーリスターン語派に改められた。
ヌーリスターン語派に属する言語
ストランドは、ヌーリスタン語派には大別して5言語が存在し、それぞれ数種類の方言をもつとしている。
主な方言にはカタヴァリ語(英語版)、カンヴィリ語(英語版)、ヴァイアラ語(英: Vai-ala)などがあり、パキスタン側のヌーリスタンではカンヴィリ語話者が大多数である。
これら方言とはダルド語派との関連が指摘されているが、言語学的というよりは地理的な関連であろう。
Askunu (Ashkun) 2,000 speakers
Kamkata-viri (Bashgali, includes the dialects Kata-vari, Kamviri & Mumviri) 24,200 speakers
Vasi-vari (Prasuni) 2,000 speakers
Tregami (Gambiri) 1,000 speakers
Waigali (Kalasha-ala) 2,000 speakers
Zemiaki 500 speakers
ヌーリスターン語派
http://ja.wikipedia.org/wiki/%E3%83%8C%E3%83%BC%E3%83%AA%E3%82%B9%E3%82%BF%E3%83%BC%E3%83%B3%E8%AA%9E%E6%B4%BE
ヌーリスターン語派(Nuristan, ヌーリスタン語派)、またはカーフィル語派(Kafir)とは、
パキスタンからアフガニスタンにかけて分布する言語のグループで、インド・ヨーロッパ語族のインド・イラン語派に属する。
比較言語学的な位置づけについては、近年ではインド・イラン語派内の独立した一グループとする見解が主流であるが、
一方で支持は少ないながらも、インド語派に含める説や、イラン語派に属するがインド語派に属するダルド語派から多大な影響を受けて変容したとする説もある。
いずれにせよ、ヌーリスターン語派の諸民族が現住地に落ち着いたのは相当の過去のことであり、またインド語派とは異なりパンジャーブ地方へは入らなかったという点に争いはない。
ヌーリスターンは1890年代までペルシア語で「不信仰者の地」を意味するカフィリスタン(英語版)と呼ばれていた。
これは当地の住民が独自の多神教(アニミズム)を信仰していたためで、言語もこれに倣ってカーフィル語派とされていた。
しかしアブドゥッラフマーン・ハーンに征服されて強制的に改宗させられ、住民の一部がイスラームを受容すると、
異教の民がイスラームの光に照らされたとしてヌーリスターン(光の地)と呼ばれるようになり、
カーフィル語派の名も差別的だとしてヌーリスターン語派に改められた。
ヌーリスターン語派に属する言語
ストランドは、ヌーリスタン語派には大別して5言語が存在し、それぞれ数種類の方言をもつとしている。
主な方言にはカタヴァリ語(英語版)、カンヴィリ語(英語版)、ヴァイアラ語(英: Vai-ala)などがあり、パキスタン側のヌーリスタンではカンヴィリ語話者が大多数である。
これら方言とはダルド語派との関連が指摘されているが、言語学的というよりは地理的な関連であろう。
Askunu (Ashkun) 2,000 speakers
Kamkata-viri (Bashgali, includes the dialects Kata-vari, Kamviri & Mumviri) 24,200 speakers
Vasi-vari (Prasuni) 2,000 speakers
Tregami (Gambiri) 1,000 speakers
Waigali (Kalasha-ala) 2,000 speakers
Zemiaki 500 speakers
968 :世界@名無史さん:2014/02/13(木) 22:21:19.40 0
ヌーリスターンの伝統宗教って、ヒンドゥー教からドラビダやムンダ由来の要素を除いたような感じなのかな
>>968どうなんっすかね
パシュトー語Pashto language
http://ja.wikipedia.org/wiki/%E3%83%91%E3%82%B7%E3%83%A5%E3%83%88%E3%83%BC%E8%AA%9E
http://upload.wikimedia.org/wikipedia/commons/thumb/8/85/Pashtun_Language_Location_Map.svg/500px-Pashtun_Language_Location_Map.svg.png
パシュトー語(パシュトーご)は、アフガニスタン、またパキスタンの西部に住むアフガン人(パシュトゥーン人)の話す言語である。
インドヨーロッパ語族のイラン語派の東語群に属す。
アフガニスタンでは、憲法によりパシュトー語が公用語と定められている。一方、パキスタンでは、国語および公用語として定められていない。
文字[編集]
アラビア文字の系統のペルシア文字に、アラビア語にもペルシア語にも存在しない音(そり舌音など)の文字を加えて改造した、パシュトー文字(英語版)と呼ばれる文字を用いる。
ナスフ体(英語版)を主に用い、ナスタアリーク体も用いられる。
北部パシュトー語 - 11,430,000人(パキスタン、アフガニスタン、アラブ首長国連邦)
ギルザイ方言 - Ghilzai (pbu-ghi)
北東パシュトー方言 - Northwestern Pakhto (pbu-nop)
Ningraharian Pashto (pbu-nin)
デュラニ方言 - Durani (pbu-dur)
中部パシュトー語 - 7,920,000人(パキスタン)
Bannuchi (pst-ban)
Waciri (pst-wac)
南部パシュトー語 - 7,590,100人(アフガニスタン、イラン、パキスタン、タジキスタン)
クエッタ・パシュトー方言 - Quetta Pashto (pbt-que)
南東パシュトー方言 - Southeastern Pashto (pbt-sou)
カンダハール・パシュトー方言 - Kandahar Pashto (pbt-kan)
南西パシュトー方言 - Southwestern Pashto (pbt-sop)
パシュトー語Pashto language
http://ja.wikipedia.org/wiki/%E3%83%91%E3%82%B7%E3%83%A5%E3%83%88%E3%83%BC%E8%AA%9E
http://upload.wikimedia.org/wikipedia/commons/thumb/8/85/Pashtun_Language_Location_Map.svg/500px-Pashtun_Language_Location_Map.svg.png
{kind=link}
パシュトー語(パシュトーご)は、アフガニスタン、またパキスタンの西部に住むアフガン人(パシュトゥーン人)の話す言語である。
インドヨーロッパ語族のイラン語派の東語群に属す。
アフガニスタンでは、憲法によりパシュトー語が公用語と定められている。一方、パキスタンでは、国語および公用語として定められていない。
文字[編集]
アラビア文字の系統のペルシア文字に、アラビア語にもペルシア語にも存在しない音(そり舌音など)の文字を加えて改造した、パシュトー文字(英語版)と呼ばれる文字を用いる。
ナスフ体(英語版)を主に用い、ナスタアリーク体も用いられる。
北部パシュトー語 - 11,430,000人(パキスタン、アフガニスタン、アラブ首長国連邦)
ギルザイ方言 - Ghilzai (pbu-ghi)
北東パシュトー方言 - Northwestern Pakhto (pbu-nop)
Ningraharian Pashto (pbu-nin)
デュラニ方言 - Durani (pbu-dur)
中部パシュトー語 - 7,920,000人(パキスタン)
Bannuchi (pst-ban)
Waciri (pst-wac)
南部パシュトー語 - 7,590,100人(アフガニスタン、イラン、パキスタン、タジキスタン)
クエッタ・パシュトー方言 - Quetta Pashto (pbt-que)
南東パシュトー方言 - Southeastern Pashto (pbt-sou)
カンダハール・パシュトー方言 - Kandahar Pashto (pbt-kan)
南西パシュトー方言 - Southwestern Pashto (pbt-sop)
>>969
History[edit]
According to 19th-century linguist James Darmesteter and modern linguist Michael M. T. Henderson, Pashto is "descended from Avestan",[9][10][11]
but Georg Morgenstierne says they are merely closely related.[47]
The word "Pashto" derives by regular phonological processes from Parsaw?- "Persian".[48]
Nonetheless, the Pashtuns are sometimes compared with the Pakhta tribes mentioned in the Rigveda (1700?1100 BC), apparently the same as a people called Pactyans,
described by the Greek historian Herodotus as living in the Achaemenid's Arachosia Satrapy as early as the 1st millennium BC.[49]
However, this comparison appears to be due mainly to the apparent, etymologically unjustified, similarity between their names.[50]
Strabo, who lived between 64 BC and 24 CE, explains that the tribes inhabiting the lands west of the Indus River were part of Ariana and to their east was India.
Since the 3rd century CE and onward, they are mostly referred to by the name "Afghan" ("Abgan")[51][52][53] and their language as "Afghani".[7]
Scholars such as Abdul Hai Habibi and others believe that the earliest modern Pashto work dates back to Amir Kror Suri in the eighth century, and they use the writings found in Pata Khazana.
However, this is disputed by several European experts due to lack of strong evidence.
Pata Khazana is a Pashto manuscript[54] claimed to be first compiled during the Hotaki dynasty (1709?1738) in Kandahar, Afghanistan.
During the 17th century Pashto poetry was becoming very popular among the Pashtuns.
Some of those who wrote poetry in Pashto are Khushal Khan Khattak, Rahman Baba, Nazo Tokhi and Ahmad Shah Durrani, founder of the modern state of Afghanistan or the Afghan Empire.
History[edit]
According to 19th-century linguist James Darmesteter and modern linguist Michael M. T. Henderson, Pashto is "descended from Avestan",[9][10][11]
but Georg Morgenstierne says they are merely closely related.[47]
The word "Pashto" derives by regular phonological processes from Parsaw?- "Persian".[48]
Nonetheless, the Pashtuns are sometimes compared with the Pakhta tribes mentioned in the Rigveda (1700?1100 BC), apparently the same as a people called Pactyans,
described by the Greek historian Herodotus as living in the Achaemenid's Arachosia Satrapy as early as the 1st millennium BC.[49]
However, this comparison appears to be due mainly to the apparent, etymologically unjustified, similarity between their names.[50]
Strabo, who lived between 64 BC and 24 CE, explains that the tribes inhabiting the lands west of the Indus River were part of Ariana and to their east was India.
Since the 3rd century CE and onward, they are mostly referred to by the name "Afghan" ("Abgan")[51][52][53] and their language as "Afghani".[7]
Scholars such as Abdul Hai Habibi and others believe that the earliest modern Pashto work dates back to Amir Kror Suri in the eighth century, and they use the writings found in Pata Khazana.
However, this is disputed by several European experts due to lack of strong evidence.
Pata Khazana is a Pashto manuscript[54] claimed to be first compiled during the Hotaki dynasty (1709?1738) in Kandahar, Afghanistan.
During the 17th century Pashto poetry was becoming very popular among the Pashtuns.
Some of those who wrote poetry in Pashto are Khushal Khan Khattak, Rahman Baba, Nazo Tokhi and Ahmad Shah Durrani, founder of the modern state of Afghanistan or the Afghan Empire.
>>969
パシュトゥーン人
http://ja.wikipedia.org/wiki/%E3%83%91%E3%82%B7%E3%83%A5%E3%83%88%E3%82%A5%E3%83%BC%E3%83%B3%E4%BA%BA
アフガニスタン(アフガーニスターン (Afgh?nist?n))は、ペルシア語・ダリー語で「アフガン人(パシュトゥーン人)の国」という意味。
北パキスタンのペシャーワルと南アフガニスタンのカンダハールは、パシュトゥーン人の伝統的な主要な中心都市である。
パシュトゥーン人は、パシュトー語またはパフトー語を話し、ダリー語とともにアフガニスタンの公用語の一つである。
歴史[編集]
紀元前2世紀後半に北方からイラン高原の東部に侵入したと伝えられており、もともとの居住地は、カンダハールの東にあるクーヒ・スライマーン山脈の近くにあったと伝承されている[1]。
10世紀頃にイスラム教を受け入れ、のちにイランのサファヴィー朝やインドのムガル帝国の支配を受けた。
その一派は18世紀初頭にサファヴィー朝に対して反乱を起こし、1722年に首都イスファハーンを陥落させるが、アフシャール朝のナーディル・シャーに敗れた。
ナーディル・シャーの死後、彼に従っていたドゥッラーニー族のパシュトゥーン人アフマド・シャー・アブダーリーはカンダハールでアフシャール朝から自立し、
アフガニスタン国家の起源となるドゥッラーニー朝を建国する。
ドゥッラーニー族が支配するアフガニスタンでは、パシュトゥーン人部族の有力者(貴族)が国家のあらゆる側面で力を持ち、国家を支配してきた。
民族の居住地域が大きく分散していないにもかかわらず、2つの国家に分割されているのは、19世紀当時にアフガン戦争によってこの地域を支配下に置いていたイギリスが、
保護国アフガニスタンと植民地インドとの境界を民族分布を考慮せずに引いたためである。
1893年の国境線(デュアランド・ライン)の画定に伴い、パシュトゥーン人の居住地域は、アフガニスタンと現在のパキスタン北西部に分かれることとなった。
パシュトゥーン人
http://ja.wikipedia.org/wiki/%E3%83%91%E3%82%B7%E3%83%A5%E3%83%88%E3%82%A5%E3%83%BC%E3%83%B3%E4%BA%BA
アフガニスタン(アフガーニスターン (Afgh?nist?n))は、ペルシア語・ダリー語で「アフガン人(パシュトゥーン人)の国」という意味。
北パキスタンのペシャーワルと南アフガニスタンのカンダハールは、パシュトゥーン人の伝統的な主要な中心都市である。
パシュトゥーン人は、パシュトー語またはパフトー語を話し、ダリー語とともにアフガニスタンの公用語の一つである。
歴史[編集]
紀元前2世紀後半に北方からイラン高原の東部に侵入したと伝えられており、もともとの居住地は、カンダハールの東にあるクーヒ・スライマーン山脈の近くにあったと伝承されている[1]。
10世紀頃にイスラム教を受け入れ、のちにイランのサファヴィー朝やインドのムガル帝国の支配を受けた。
その一派は18世紀初頭にサファヴィー朝に対して反乱を起こし、1722年に首都イスファハーンを陥落させるが、アフシャール朝のナーディル・シャーに敗れた。
ナーディル・シャーの死後、彼に従っていたドゥッラーニー族のパシュトゥーン人アフマド・シャー・アブダーリーはカンダハールでアフシャール朝から自立し、
アフガニスタン国家の起源となるドゥッラーニー朝を建国する。
ドゥッラーニー族が支配するアフガニスタンでは、パシュトゥーン人部族の有力者(貴族)が国家のあらゆる側面で力を持ち、国家を支配してきた。
民族の居住地域が大きく分散していないにもかかわらず、2つの国家に分割されているのは、19世紀当時にアフガン戦争によってこの地域を支配下に置いていたイギリスが、
保護国アフガニスタンと植民地インドとの境界を民族分布を考慮せずに引いたためである。
1893年の国境線(デュアランド・ライン)の画定に伴い、パシュトゥーン人の居住地域は、アフガニスタンと現在のパキスタン北西部に分かれることとなった。
>>967
Kafiristan
http://en.wikipedia.org/wiki/Kafiristan
http://upload.wikimedia.org/wikipedia/commons/4/48/AfghanistanNurestan.png
History
Ancient
Further information: History of Afghanistan
Ancient Kapi?a Janapada, located south-east of the Hindukush, included and is related to Kafiristan.[5]
The Chinese pilgrim Hiuen Tsang who visited Kapisa in 644 AD calls it Kai-pi-shi(h).[6]
Hiuen Tsang describes Kai-pi-shi[7] as a flourishing kingdom ruled by a Buddhist Kshatriya king holding sway over ten neighbouring states including Lampaka, Nagarahara, Gandhara and Bannu.
Until the 9th century AD, Kapi?i remained the second capital of the Shahi dynasty of Kabul.
Kapi?a was known for goats and their skin.[8] Hiuen Tsang talks of Shen breed of horses from Kapi?a (Kai-pi-shi).
There is also a reference to Chinese emperor Tai-Tsung being presented with excellent breed of horses in 637 AD by an envoy from Chi-pin (Kapisa).[9]
Further evidence from Hiuen Tsang shows that Kai-pi-shi produced all kind of cereals, many kinds of fruits, and a scented root called Yu-kin, probably khus or vetiver.
The people used woollen and fur clothes and gold,[10][11] silver and copper coins.
Objects of merchandise from all parts were found here.[12]
Kafiristan
http://en.wikipedia.org/wiki/Kafiristan
http://upload.wikimedia.org/wikipedia/commons/4/48/AfghanistanNurestan.png
{kind=link}
History
Ancient
Further information: History of Afghanistan
Ancient Kapi?a Janapada, located south-east of the Hindukush, included and is related to Kafiristan.[5]
The Chinese pilgrim Hiuen Tsang who visited Kapisa in 644 AD calls it Kai-pi-shi(h).[6]
Hiuen Tsang describes Kai-pi-shi[7] as a flourishing kingdom ruled by a Buddhist Kshatriya king holding sway over ten neighbouring states including Lampaka, Nagarahara, Gandhara and Bannu.
Until the 9th century AD, Kapi?i remained the second capital of the Shahi dynasty of Kabul.
Kapi?a was known for goats and their skin.[8] Hiuen Tsang talks of Shen breed of horses from Kapi?a (Kai-pi-shi).
There is also a reference to Chinese emperor Tai-Tsung being presented with excellent breed of horses in 637 AD by an envoy from Chi-pin (Kapisa).[9]
Further evidence from Hiuen Tsang shows that Kai-pi-shi produced all kind of cereals, many kinds of fruits, and a scented root called Yu-kin, probably khus or vetiver.
The people used woollen and fur clothes and gold,[10][11] silver and copper coins.
Objects of merchandise from all parts were found here.[12]
>>968
ヌーリスターン人
http://ja.wikipedia.org/wiki/%E3%83%8C%E3%83%BC%E3%83%AA%E3%82%B9%E3%82%BF%E3%83%BC%E3%83%B3%E4%BA%BA
ヌーリスターン人(ヌーリスターンじん、Nuristani)は、中央アジア・南アジアのアフガニスタンを中心に居住する民族である。ヌーリスタン人やヌリスタニ人とも呼ばれる。
http://en.wikipedia.org/wiki/File:Girl_in_a_Kabul_orphanage,_01-07-2002.jpg
アフガニスタン北東部のヌーリスターン州に住み、一部はパキスタンの北西辺境州 のチトラル地区にも住む。
金髪、紺碧目で色白という北ヨーロッパ人と同じような外見を持ち、特に子ども時代はその傾向が強い。
そのため、一説には当時この地を通った、ギリシャのアレキサンダー大王の軍隊の末裔であるという説もある。
言語はインドヨーロッパ語族イラン語派のヌーリスターン語派であるが、第二言語としてダリー語やペルシャ語、パシュトー語も話す。
ダリー語(ファルシー語)
http://ja.wikipedia.org/wiki/%E3%83%80%E3%83%AA%E3%83%BC%E8%AA%9E
ダリー語はアフガニスタンの公用語の一つ。タジク語と同じく基本的にはペルシア語であるが、発音・語彙を中心にいくらかの違いがある。
発音は、ペルシア語のe, oがダリー語ではi, uになるなど、ほぼ組織的な対応関係がある。
語彙の大半はペルシア語と同じだが、ダリー語にはパシュトー語やウルドゥー語からの借用語が多い。
また、西欧語からの新しい借用語については、ペルシア語ではフランス語由来の語が多いのに対し、ダリー語では英語由来のものが多い。
うーん、なにかの遺跡が思い出せない、、、
ヌーリスターン人
http://ja.wikipedia.org/wiki/%E3%83%8C%E3%83%BC%E3%83%AA%E3%82%B9%E3%82%BF%E3%83%BC%E3%83%B3%E4%BA%BA
ヌーリスターン人(ヌーリスターンじん、Nuristani)は、中央アジア・南アジアのアフガニスタンを中心に居住する民族である。ヌーリスタン人やヌリスタニ人とも呼ばれる。
http://en.wikipedia.org/wiki/File:Girl_in_a_Kabul_orphanage,_01-07-2002.jpg
{kind=link}
アフガニスタン北東部のヌーリスターン州に住み、一部はパキスタンの北西辺境州 のチトラル地区にも住む。
金髪、紺碧目で色白という北ヨーロッパ人と同じような外見を持ち、特に子ども時代はその傾向が強い。
そのため、一説には当時この地を通った、ギリシャのアレキサンダー大王の軍隊の末裔であるという説もある。
言語はインドヨーロッパ語族イラン語派のヌーリスターン語派であるが、第二言語としてダリー語やペルシャ語、パシュトー語も話す。
ダリー語(ファルシー語)
http://ja.wikipedia.org/wiki/%E3%83%80%E3%83%AA%E3%83%BC%E8%AA%9E
ダリー語はアフガニスタンの公用語の一つ。タジク語と同じく基本的にはペルシア語であるが、発音・語彙を中心にいくらかの違いがある。
発音は、ペルシア語のe, oがダリー語ではi, uになるなど、ほぼ組織的な対応関係がある。
語彙の大半はペルシア語と同じだが、ダリー語にはパシュトー語やウルドゥー語からの借用語が多い。
また、西欧語からの新しい借用語については、ペルシア語ではフランス語由来の語が多いのに対し、ダリー語では英語由来のものが多い。
うーん、なにかの遺跡が思い出せない、、、
さて、スレも残り少ないので雑感を
>>600 ミステリートライアングル
>>574 一部判明したが気になる部分が残ってる
>>557 不明、検証したい
>>611
どっかでNGSDの話を見かけた
そしてワスレタww
>>687 たぶん、まだ頑張れない
>>720>>724
これはやることある
>>768 家畜は継続
いつもの詳しい人に教えてもらいたい
>>801よりは>>806
>>791 レムノス島とロドス島は、クレタも含めて検証要
>>849 とりあえず落ちついて嫁>>858
>>862 ここらへんは他のも含めて年代をきっちりさせたい
>>874 結果が気になる。 たぶんドイツ、デンマークのと一緒に出てくるとみた。
>>610 そろそろ見れるかと思う
とりあえず、「海の民」だけほったらかしてる
>>600 ミステリートライアングル
>>574 一部判明したが気になる部分が残ってる
>>557 不明、検証したい
>>611
どっかでNGSDの話を見かけた
そしてワスレタww
>>687 たぶん、まだ頑張れない
>>720>>724
これはやることある
>>768 家畜は継続
いつもの詳しい人に教えてもらいたい
>>801よりは>>806
>>791 レムノス島とロドス島は、クレタも含めて検証要
>>849 とりあえず落ちついて嫁>>858
>>862 ここらへんは他のも含めて年代をきっちりさせたい
>>874 結果が気になる。 たぶんドイツ、デンマークのと一緒に出てくるとみた。
>>610 そろそろ見れるかと思う
とりあえず、「海の民」だけほったらかしてる
>>968
るつぼですね
http://www.hansen-jp.com/208amabe.htm
エスニック諸集団
このような事情から、アフガニスタンでは、南東部を中心に居住する征服者パシュトゥーン人が人口の約四〇%を占めるにすぎず、
そのほかに、
バダフシャーン地方にタジク人、
ヘラート地方とスィースターン地方にファールスィー人(ペルシア人)、
バクトリア地方にウズベク人、トルクメニスタンとの国境付近にトルクメン人、
ヒンドゥークシュ山脈の東部ハザーラジャート地方にハザーラ人、
その西部ゴール地方にアイマク(チャハール)人などが存在する。
パシュトゥーン人は一八世紀以後、スィースターン北部のファラーフ、スィンダンドやバクトリア地方にも移住し、分布を複雑なものにしている。
以上が七大エスニック集団(民族意識に乏しく、民族とはいえない)であるが、このほかに、マイノリティ集団が少なくない。
以上のエスニック集団の中で、パシュトゥーン人が話すパシュト語はイラン系であるが、ペルシア語とは大きく異なる。
タジク人、ファールスィー人、ハザーラ人、アイマク人、キズィルバーシュ人はいずれもペルシア語(ダリー語)やその方言を話すが、
ハザーラ人のペルシア語はモンゴル語の単語を含み、
アイマク人のペルシア語は、トルコ系の単語が著しく多い。
また、ウズベク語とトルクメン語は中央アジアのトルコ系言語である。
るつぼですね
http://www.hansen-jp.com/208amabe.htm
エスニック諸集団
このような事情から、アフガニスタンでは、南東部を中心に居住する征服者パシュトゥーン人が人口の約四〇%を占めるにすぎず、
そのほかに、
バダフシャーン地方にタジク人、
ヘラート地方とスィースターン地方にファールスィー人(ペルシア人)、
バクトリア地方にウズベク人、トルクメニスタンとの国境付近にトルクメン人、
ヒンドゥークシュ山脈の東部ハザーラジャート地方にハザーラ人、
その西部ゴール地方にアイマク(チャハール)人などが存在する。
パシュトゥーン人は一八世紀以後、スィースターン北部のファラーフ、スィンダンドやバクトリア地方にも移住し、分布を複雑なものにしている。
以上が七大エスニック集団(民族意識に乏しく、民族とはいえない)であるが、このほかに、マイノリティ集団が少なくない。
以上のエスニック集団の中で、パシュトゥーン人が話すパシュト語はイラン系であるが、ペルシア語とは大きく異なる。
タジク人、ファールスィー人、ハザーラ人、アイマク人、キズィルバーシュ人はいずれもペルシア語(ダリー語)やその方言を話すが、
ハザーラ人のペルシア語はモンゴル語の単語を含み、
アイマク人のペルシア語は、トルコ系の単語が著しく多い。
また、ウズベク語とトルクメン語は中央アジアのトルコ系言語である。
クローヴィス人のゲノム
http://sicambre.at.webry.info/201402/article_15.html
この研究では、アメリカ合衆国モンタナ州西部にあるクローヴィス文化のアンジック遺跡で発見された男児(Anzick-1)のゲノムが解析され、他のゲノムと比較されています。
その男児の年代は、放射性炭素年代測定法で10705±35年前頃(暦年代では12707〜12556年前頃)となります。
この男児のゲノムが現代人や更新世シベリア人と比較されました。その結果、“Anzick-1”の属していた集団は、
現代のアメリカ大陸先住民の約8割の直接的祖先であり、残りの2割も、他のどの集団よりも“Anzick-1”集団に近い関係にあることが明らかになりました。
また、“Anzick-1”はシベリアのバイカル湖近くのマリタ遺跡で発見された24000年前頃の少年と、遺伝子の1/3を共有しており、
これは現在の全アメリカ先住民も同様です。
このことから、アメリカ先住民の祖先の一部たる東アジアの集団と、更新世のマリタの集団とが出会ったのは、
クローヴィス文化以前だろう、と推測されています。以下は『ネイチャー』の日本語サイトからの引用です。
参考文献:
Rasmussen M. et al.(2014): The genome of a Late Pleistocene human from a Clovis burial site in western Montana. Nature, 506, 7487, 225?229.
http://dx.doi.org/10.1038/nature13025
http://sicambre.at.webry.info/201402/article_15.html
この研究では、アメリカ合衆国モンタナ州西部にあるクローヴィス文化のアンジック遺跡で発見された男児(Anzick-1)のゲノムが解析され、他のゲノムと比較されています。
その男児の年代は、放射性炭素年代測定法で10705±35年前頃(暦年代では12707〜12556年前頃)となります。
この男児のゲノムが現代人や更新世シベリア人と比較されました。その結果、“Anzick-1”の属していた集団は、
現代のアメリカ大陸先住民の約8割の直接的祖先であり、残りの2割も、他のどの集団よりも“Anzick-1”集団に近い関係にあることが明らかになりました。
また、“Anzick-1”はシベリアのバイカル湖近くのマリタ遺跡で発見された24000年前頃の少年と、遺伝子の1/3を共有しており、
これは現在の全アメリカ先住民も同様です。
このことから、アメリカ先住民の祖先の一部たる東アジアの集団と、更新世のマリタの集団とが出会ったのは、
クローヴィス文化以前だろう、と推測されています。以下は『ネイチャー』の日本語サイトからの引用です。
参考文献:
Rasmussen M. et al.(2014): The genome of a Late Pleistocene human from a Clovis burial site in western Montana. Nature, 506, 7487, 225?229.
http://dx.doi.org/10.1038/nature13025
>>976
古代ゲノム学:モンタナ州西部にあるクローヴィス人の墓地遺跡で発掘された更新世後期のヒトのゲノム
古代ゲノム学:古代ゲノムからアメリカ先住民の祖先が明らかに
クローヴィス文化は、約1万3000年前の北米に広く分布した古代の文化で、
クローヴィス尖頭器として知られる槍の刃など、独特な石器が特徴である。
ただ、これらの石器をどのような人々が作ったのかについては、わずかな情報しかなく、推測の域を出なかった。
今回、古代の北米にいたアメリカ先住民個体のゲノム塩基配列が初めて解読され、さらなる研究を進めるための情報が得られた。
このゲノムは米国モンタナ州にあるクローヴィス文化のアンジック遺跡の墓地で発掘された幼い男児(Anzick-1)のものである。
この部分骨格は約1万2600年前に埋葬されたもので、黄土を塗った多数の石器と一緒に出土した。
このゲノムは、現代のアメリカ先住民の祖先に当たる1つの集団に属している。
また、このゲノムと南北アメリカのさまざまな先住民集団との類縁関係は全て、他大陸のどの集団とよりも近かった。
これらの知見は、クローヴィス人がヨーロッパから移動してきたという仮説を否定するもので、クローヴィス人よりも数千年前にアメリカ大陸にヒトが居住していたという説と一致し、
現代のアメリカ先住民が、アメリカ大陸に最初にうまく定住した人々の子孫であることを示している。
http://www.nature.com/nature/journal/v506/n7487/fig_tab/nature13025_ft.html
地図http://www.nature.com/nature/journal/v506/n7487/fig_tab/nature13025_F1.html
分布http://www.nature.com/nature/journal/v506/n7487/fig_tab/nature13025_F2.html
分岐http://www.nature.com/nature/journal/v506/n7487/fig_tab/nature13025_F3.html
古代ゲノム学:モンタナ州西部にあるクローヴィス人の墓地遺跡で発掘された更新世後期のヒトのゲノム
古代ゲノム学:古代ゲノムからアメリカ先住民の祖先が明らかに
クローヴィス文化は、約1万3000年前の北米に広く分布した古代の文化で、
クローヴィス尖頭器として知られる槍の刃など、独特な石器が特徴である。
ただ、これらの石器をどのような人々が作ったのかについては、わずかな情報しかなく、推測の域を出なかった。
今回、古代の北米にいたアメリカ先住民個体のゲノム塩基配列が初めて解読され、さらなる研究を進めるための情報が得られた。
このゲノムは米国モンタナ州にあるクローヴィス文化のアンジック遺跡の墓地で発掘された幼い男児(Anzick-1)のものである。
この部分骨格は約1万2600年前に埋葬されたもので、黄土を塗った多数の石器と一緒に出土した。
このゲノムは、現代のアメリカ先住民の祖先に当たる1つの集団に属している。
また、このゲノムと南北アメリカのさまざまな先住民集団との類縁関係は全て、他大陸のどの集団とよりも近かった。
これらの知見は、クローヴィス人がヨーロッパから移動してきたという仮説を否定するもので、クローヴィス人よりも数千年前にアメリカ大陸にヒトが居住していたという説と一致し、
現代のアメリカ先住民が、アメリカ大陸に最初にうまく定住した人々の子孫であることを示している。
http://www.nature.com/nature/journal/v506/n7487/fig_tab/nature13025_ft.html
地図http://www.nature.com/nature/journal/v506/n7487/fig_tab/nature13025_F1.html
分布http://www.nature.com/nature/journal/v506/n7487/fig_tab/nature13025_F2.html
分岐http://www.nature.com/nature/journal/v506/n7487/fig_tab/nature13025_F3.html
>>977
mtDNA and Y-chromosome subtrees.
http://www.nature.com/nature/journal/v506/n7487/fig_tab/nature13025_SF2.html
http://www.nature.com/nature/journal/v506/n7487/images/nature13025-sf2.jpg
a, Schematic phylogenetic tree of mtDNA haplogroup D4h3 and its sub-branch D4h3a.
Mutations from the root of haplogroup D4h are specified only for haplogroup D4h3a lineage; diagnostic mutations are shown only for defined sub-branches on solid lines.
The haplotypes of Anzick-1, identical with the root haplotype of D4h3a,
and an ancient full sequence from the northwestern coast of North America (Ancient939), are indicated in red.
Insertions are indicated with ‘.’ followed by a number of inserted nucleotides (X if not specified), deletions are indicated with ‘d’ and back mutations to ancestral state with ‘!’.
The geographical spread of sub-branches of haplogroup D4h is shown with different colours specified in figure legend.
b, Placement of Anzick-1 within the Y-chromosome phylogeny.
Anzick-1 (circled) represents Y-chromosome haplogroup Q-L54*(xM3) (blue), which is offset by haplogroup Q-M3 (dark blue).
The lineage carried by the ancient Saqqaq Palaeo-Eskimo (light blue) constitutes an outgroup to Q-L54.
Each branch is labelled by an index and the number of transversion SNPs assigned to the branch (in brackets).
Terminal taxa (individuals) are also labelled by population, ID and haplogroup.
Branches 21 and 25 represent the most recent shared ancestry between Anzick-1 and other members of the sample.
Branch 19 is considerably shorter than neighbouring branches, which have had an additional ~12,600 years to accumulate mutations.
みれる?
http://www.nature.com/nature/journal/v506/n7487/images/nature13025-f1.jpg
http://www.nature.com/nature/journal/v506/n7487/images/nature13025-f2.jpg
http://www.nature.com/nature/journal/v506/n7487/images/nature13025-f3.jpg
mtDNA and Y-chromosome subtrees.
http://www.nature.com/nature/journal/v506/n7487/fig_tab/nature13025_SF2.html
http://www.nature.com/nature/journal/v506/n7487/images/nature13025-sf2.jpg
{kind=link}
a, Schematic phylogenetic tree of mtDNA haplogroup D4h3 and its sub-branch D4h3a.
Mutations from the root of haplogroup D4h are specified only for haplogroup D4h3a lineage; diagnostic mutations are shown only for defined sub-branches on solid lines.
The haplotypes of Anzick-1, identical with the root haplotype of D4h3a,
and an ancient full sequence from the northwestern coast of North America (Ancient939), are indicated in red.
Insertions are indicated with ‘.’ followed by a number of inserted nucleotides (X if not specified), deletions are indicated with ‘d’ and back mutations to ancestral state with ‘!’.
The geographical spread of sub-branches of haplogroup D4h is shown with different colours specified in figure legend.
b, Placement of Anzick-1 within the Y-chromosome phylogeny.
Anzick-1 (circled) represents Y-chromosome haplogroup Q-L54*(xM3) (blue), which is offset by haplogroup Q-M3 (dark blue).
The lineage carried by the ancient Saqqaq Palaeo-Eskimo (light blue) constitutes an outgroup to Q-L54.
Each branch is labelled by an index and the number of transversion SNPs assigned to the branch (in brackets).
Terminal taxa (individuals) are also labelled by population, ID and haplogroup.
Branches 21 and 25 represent the most recent shared ancestry between Anzick-1 and other members of the sample.
Branch 19 is considerably shorter than neighbouring branches, which have had an additional ~12,600 years to accumulate mutations.
みれる?
http://www.nature.com/nature/journal/v506/n7487/images/nature13025-f1.jpg
{kind=link}
http://www.nature.com/nature/journal/v506/n7487/images/nature13025-f2.jpg
{kind=link}
http://www.nature.com/nature/journal/v506/n7487/images/nature13025-f3.jpg
{kind=link}
>>600
なんで見てなかったんだろうww
まじワロス
Table SI 14
Pigmentation prediction in the MA-1 individual contrasted with the Tyrolean Iceman
The method is based on assessing a total 124 SNPs associated with pigmentation traits
Phenotype
MA-1 (17 SNPs)
Iceman (24 SNPs)
darker skin vs fairer skin
blue eyes vs non-blue eyes
brown eyes vs non-brown eyes
green or blue eyes vs brown or black eyes
red hair vs non-red hair
freckles vs non-freckles
blond hair vs non-blond hair
brown hair vs non-brown hair
lighter brown or blond hair vs darker brown or black hair
blond or red hair vs brown hair
lighter brown hair vs darker brown hair
blond or red hair vs non-blond or non-red hair
なんで見てなかったんだろうww
まじワロス
Table SI 14
Pigmentation prediction in the MA-1 individual contrasted with the Tyrolean Iceman
The method is based on assessing a total 124 SNPs associated with pigmentation traits
Phenotype
MA-1 (17 SNPs)
Iceman (24 SNPs)
darker skin vs fairer skin
blue eyes vs non-blue eyes
brown eyes vs non-brown eyes
green or blue eyes vs brown or black eyes
red hair vs non-red hair
freckles vs non-freckles
blond hair vs non-blond hair
brown hair vs non-brown hair
lighter brown or blond hair vs darker brown or black hair
blond or red hair vs brown hair
lighter brown hair vs darker brown hair
blond or red hair vs non-blond or non-red hair
>>979
MA-1 (17 SNPs)
darker skin (6/9)
Non-blue eyes (4/4)
brown eyes (2/3)
brown or black eyes (2/2)
-
freckles (1/1)
Non-blond hair (1/1)
brown hair (5/7)
darker brown or black hair (2/3)
brown hair (7/7)
lighter brown hair (2/2)
-
Iceman (24 SNPs)
fairer skin (8/15)
blue eyes (7/9)
non-brown eyes (5/8)
brown or black eyes (3/3)
red hair (2/3)
freckles (2/3)
blond hair (2/3)
brown hair (8/11)
lighter brown or blond hair (2/3)
brown hair (11/13)
lighter brown hair (2/3)
blond or red hair (2/3)
MA-1 (17 SNPs)
darker skin (6/9)
Non-blue eyes (4/4)
brown eyes (2/3)
brown or black eyes (2/2)
-
freckles (1/1)
Non-blond hair (1/1)
brown hair (5/7)
darker brown or black hair (2/3)
brown hair (7/7)
lighter brown hair (2/2)
-
Iceman (24 SNPs)
fairer skin (8/15)
blue eyes (7/9)
non-brown eyes (5/8)
brown or black eyes (3/3)
red hair (2/3)
freckles (2/3)
blond hair (2/3)
brown hair (8/11)
lighter brown or blond hair (2/3)
brown hair (11/13)
lighter brown hair (2/3)
blond or red hair (2/3)
>>610
Table S2
Population(X) n Language Social/ caste
Paniya 5 Dravidian Tribal
Palliyar 5 Dravidian Tribal
Kattunayakan 5 Dravidian Tribal
Palliyar^a 5 Dravidian Lower caste
Madiga 13 Dravidian Lower caste
Mala 13 Dravidian Lower caste
Adi-Dravidar 5 Dravidian Lower caste
Hakkipikki^a 4 Dravidian Tribal
Vedda 4 Indo-European Tribal
Kamsali 4 Dravidian Lower caste
Chenchu^a 4 Dravidian Tribal
Chamar^a 10 Indo-European Tribal
Chenchu 6 Dravidian Tribal
Bhil 17 Indo-European Tribal
Kallar 5 Dravidian Lower caste
Kallar^a 8 Dravidian Tribal
Vysya 14 Dravidian Middle caste
Malai Kuravar 5 Dravidian Tribal
Satnami 3 Indo-European Lower caste
Kuruchiyan 5 Dravidian Tribal
Dushadh^a 7 Indo-European Lower caste
Scheduled
caste/tribe^a 6 Dravidian Lower caste
Table S2
Population(X) n Language Social/ caste
Paniya 5 Dravidian Tribal
Palliyar 5 Dravidian Tribal
Kattunayakan 5 Dravidian Tribal
Palliyar^a 5 Dravidian Lower caste
Madiga 13 Dravidian Lower caste
Mala 13 Dravidian Lower caste
Adi-Dravidar 5 Dravidian Lower caste
Hakkipikki^a 4 Dravidian Tribal
Vedda 4 Indo-European Tribal
Kamsali 4 Dravidian Lower caste
Chenchu^a 4 Dravidian Tribal
Chamar^a 10 Indo-European Tribal
Chenchu 6 Dravidian Tribal
Bhil 17 Indo-European Tribal
Kallar 5 Dravidian Lower caste
Kallar^a 8 Dravidian Tribal
Vysya 14 Dravidian Middle caste
Malai Kuravar 5 Dravidian Tribal
Satnami 3 Indo-European Lower caste
Kuruchiyan 5 Dravidian Tribal
Dushadh^a 7 Indo-European Lower caste
Scheduled
caste/tribe^a 6 Dravidian Lower caste
>>981
Population(X) n Language Social/ caste
Mali 5 Dravidian Lower caste
Minicoy 4 Indo-European Lower caste
Gounder 5 Dravidian Middle caste
Lodi 5 Indo-European Lower caste
Naidu 4 Dravidian Upper caste
Velama 4 Dravidian Upper caste
Velama^a 9 Dravidian Upper caste
Narikkuravar 5 Dravidian Tribal
Tharu 5 Indo-European Tribal
Dharkar^a 11 Indo-European Nomadic group
Kanjar^a 8 Indo-European Nomadic group
Muslim^a 5 Indo-European Religious group
Srivastava 2 Indo-European Upper caste
Jain 5 Indo-European Religious group
Meghawal 5 Indo-European Lower caste
Kshatriya^a 7 Indo-European Upper caste
Vaish 4 Indo-European Upper caste
Brahmin^a 8 Indo-European Upper caste
Kshatriy^a 15 Indo-European Upper caste
Brahmin 10 Indo-European Upper caste
Sindhi^b 10 Indo-European Urban group
Kashmiri Pandit 15 Indo-European Upper caste
Pathan^b 15 Indo-European Urban group
Population(X) n Language Social/ caste
Mali 5 Dravidian Lower caste
Minicoy 4 Indo-European Lower caste
Gounder 5 Dravidian Middle caste
Lodi 5 Indo-European Lower caste
Naidu 4 Dravidian Upper caste
Velama 4 Dravidian Upper caste
Velama^a 9 Dravidian Upper caste
Narikkuravar 5 Dravidian Tribal
Tharu 5 Indo-European Tribal
Dharkar^a 11 Indo-European Nomadic group
Kanjar^a 8 Indo-European Nomadic group
Muslim^a 5 Indo-European Religious group
Srivastava 2 Indo-European Upper caste
Jain 5 Indo-European Religious group
Meghawal 5 Indo-European Lower caste
Kshatriya^a 7 Indo-European Upper caste
Vaish 4 Indo-European Upper caste
Brahmin^a 8 Indo-European Upper caste
Kshatriy^a 15 Indo-European Upper caste
Brahmin 10 Indo-European Upper caste
Sindhi^b 10 Indo-European Urban group
Kashmiri Pandit 15 Indo-European Upper caste
Pathan^b 15 Indo-European Urban group
>>982
Palliyar^a 5 Dravidian Lower caste Cypriot > Abhkasian (0.4) > Georgian (0.5)
Brahmina 8 Indo-European Upper caste Tuscan > Lezgin (0.0) > Georgian (0.1)
他はじょーじ
We compute D(Onge, X; YRI, Y) where X is an Indian group shown above and Y is a West Eurasian group chosen from a panel of 43 groups
including Europeans, Central Asians, Middle Easterners and Caucasian populations.
We display the results for the population Y with the highest D-statistic mean,
the 2nd highest D-statistic mean (Z-score for the difference between highest and 2nd highest group),
and the 3nd highest D-statistic mean (Z-score for the difference between the highest and 3nd highest). We consider
|Z| > 3 to be statistically significant. a
indicates samples from Metspalu et al (2011) and b
indicates samples from HGDP.
Palliyar^a 5 Dravidian Lower caste Cypriot > Abhkasian (0.4) > Georgian (0.5)
Brahmina 8 Indo-European Upper caste Tuscan > Lezgin (0.0) > Georgian (0.1)
他はじょーじ
We compute D(Onge, X; YRI, Y) where X is an Indian group shown above and Y is a West Eurasian group chosen from a panel of 43 groups
including Europeans, Central Asians, Middle Easterners and Caucasian populations.
We display the results for the population Y with the highest D-statistic mean,
the 2nd highest D-statistic mean (Z-score for the difference between highest and 2nd highest group),
and the 3nd highest D-statistic mean (Z-score for the difference between the highest and 3nd highest). We consider
|Z| > 3 to be statistically significant. a
indicates samples from Metspalu et al (2011) and b
indicates samples from HGDP.
>>981
http://en.wikipedia.org/wiki/List_of_Scheduled_Tribes_in_India
>>983
Palliyar people
http://www.joshuaproject.net/people-profile.php?rog3=IN&peo3=17889
Religion
Buddhism 0.00 %
Christianity 0.77 % (Evangelical: Data not available)
Ethnic Religions 0.00 %
Hinduism 99.23 %
Islam 0.00 %
Non-Religious 0.00 %
Other / Small 0.00 %
Unknown 0.00 %
http://en.wikipedia.org/wiki/Brahmin
Brahmin (/?br??m?n/; also called Brahmana; from the Sanskrit br?hma?a ????????) are traditional Hindu societies of India, Nepal and The Far East.
Brahman, Brahmin, and Brahma have different meanings.
Brahman refers to the Supreme Self. Brahmin (or Brahmana) refers to an individual belonging to the Hindu priest, artists, teachers, technicians class (varna or pillar of the society)
and also to an individual belonging to the Brahmin tribe/caste into which an individual is born; while the word Brahma refers to the creative aspect of the universal consciousness or God.
Because the priest / Acharya is knowledgeable about Brahma (the God), and is responsible for religious rituals in temples and homes and is a person authorized after rigorous training in vedas (sacred texts of knowledge)
and religious rituals to provide advice and impart knowledge of God to members of the society and assist in attainment of moksha,
the liberation from life cycle; the priest / Acharya class is called "Brahmin varna." The English word brahmin is an anglicized form of the Sanskrit word Br?hmana.
わからんww
http://en.wikipedia.org/wiki/List_of_Scheduled_Tribes_in_India
>>983
Palliyar people
http://www.joshuaproject.net/people-profile.php?rog3=IN&peo3=17889
Religion
Buddhism 0.00 %
Christianity 0.77 % (Evangelical: Data not available)
Ethnic Religions 0.00 %
Hinduism 99.23 %
Islam 0.00 %
Non-Religious 0.00 %
Other / Small 0.00 %
Unknown 0.00 %
http://en.wikipedia.org/wiki/Brahmin
Brahmin (/?br??m?n/; also called Brahmana; from the Sanskrit br?hma?a ????????) are traditional Hindu societies of India, Nepal and The Far East.
Brahman, Brahmin, and Brahma have different meanings.
Brahman refers to the Supreme Self. Brahmin (or Brahmana) refers to an individual belonging to the Hindu priest, artists, teachers, technicians class (varna or pillar of the society)
and also to an individual belonging to the Brahmin tribe/caste into which an individual is born; while the word Brahma refers to the creative aspect of the universal consciousness or God.
Because the priest / Acharya is knowledgeable about Brahma (the God), and is responsible for religious rituals in temples and homes and is a person authorized after rigorous training in vedas (sacred texts of knowledge)
and religious rituals to provide advice and impart knowledge of God to members of the society and assist in attainment of moksha,
the liberation from life cycle; the priest / Acharya class is called "Brahmin varna." The English word brahmin is an anglicized form of the Sanskrit word Br?hmana.
わからんww
>>983
http://en.wikipedia.org/wiki/Lezgian_people
he Lezgians (Lezgian: лезгияр, lezgiyar, Russian: лезгины, lezginy; also called Lezgins, Lezgi, Lezgis, Lezgs, Lezgin)
are an ethnic group living predominantly in southern Dagestan and northeastern Azerbaijan and who speak the Lezgian language.
http://en.wikipedia.org/wiki/File:Lezgin_map.png
History[edit]In the 4th century BC, the numerous tribes speaking Lezgic languages, which is part of the Nakh-Dagestan family of languages,
united in a union of 26 tribes, formed in the Eastern Caucasus state of Caucasian Albania, which existed before the 8th century BC.
Under the influence of foreign invaders Caucasian Albania was divided into several areas - Lakzi, Shirvan, etc.
http://en.wikipedia.org/wiki/Lezgian_people
he Lezgians (Lezgian: лезгияр, lezgiyar, Russian: лезгины, lezginy; also called Lezgins, Lezgi, Lezgis, Lezgs, Lezgin)
are an ethnic group living predominantly in southern Dagestan and northeastern Azerbaijan and who speak the Lezgian language.
http://en.wikipedia.org/wiki/File:Lezgin_map.png
{kind=link}
History[edit]In the 4th century BC, the numerous tribes speaking Lezgic languages, which is part of the Nakh-Dagestan family of languages,
united in a union of 26 tribes, formed in the Eastern Caucasus state of Caucasian Albania, which existed before the 8th century BC.
Under the influence of foreign invaders Caucasian Albania was divided into several areas - Lakzi, Shirvan, etc.
>>968
詳しくわかんないけど>>972に
Hiuen Tsang describes Kai-pi-shi[7] as a flourishing kingdom ruled by a Buddhist Kshatriya king
holding sway over ten neighbouring states including Lampaka, Nagarahara, Gandhara and Bannu.
ってあるよ。
個人的にはゾロアスター教は色々と調べたけど、
ヒンドゥー教はあんまりなじみがない。
詳しくわかんないけど>>972に
Hiuen Tsang describes Kai-pi-shi[7] as a flourishing kingdom ruled by a Buddhist Kshatriya king
holding sway over ten neighbouring states including Lampaka, Nagarahara, Gandhara and Bannu.
ってあるよ。
個人的にはゾロアスター教は色々と調べたけど、
ヒンドゥー教はあんまりなじみがない。
>>610 上の方
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2842210/bin/nihms137159f1.jpg
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2842210/bin/nihms137159f3.jpg
Indian Cline group Samples Z-score from 3 Population Test for mixture % ANI ancestry
Mala 3 -2.5 38.80%
Madiga 4 -2.7 40.60%
Chenchu 6 31.3 40.70%
Bhil 7 -10.6 42.90%
Satnami 3 -5.6 43.00%
Kurumba 6 -12.6 43.20%
Kamsali 3 -6.5 44.50%
Vysya 5 5.4 46.20%
Lodi 5 -8.9 49.90%
Naidu 4 -3.3 50.10%
Tharu 5 -20.6 51.00%
Velama 4 -3.2 54.70%
Srivastava 2 -7.5 56.40%
Meghawal 5 -13.3 60.30%
Vaish 4 -22 62.60%
Kashmiri
Pandit 5 -20.6 70.60%
Sindhi 10 -26.3 73.70%
Pathan 15 -34.3 76.90%
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2842210/bin/nihms137159f1.jpg
{kind=link}
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2842210/bin/nihms137159f3.jpg
{kind=link}
Indian Cline group Samples Z-score from 3 Population Test for mixture % ANI ancestry
Mala 3 -2.5 38.80%
Madiga 4 -2.7 40.60%
Chenchu 6 31.3 40.70%
Bhil 7 -10.6 42.90%
Satnami 3 -5.6 43.00%
Kurumba 6 -12.6 43.20%
Kamsali 3 -6.5 44.50%
Vysya 5 5.4 46.20%
Lodi 5 -8.9 49.90%
Naidu 4 -3.3 50.10%
Tharu 5 -20.6 51.00%
Velama 4 -3.2 54.70%
Srivastava 2 -7.5 56.40%
Meghawal 5 -13.3 60.30%
Vaish 4 -22 62.60%
Kashmiri
Pandit 5 -20.6 70.60%
Sindhi 10 -26.3 73.70%
Pathan 15 -34.3 76.90%
ヨーロッパにおけるジプシーの放浪生活は1500年前に始まった、DNA調査で明らかに
http://jp.sciencenewsline.com/articles/2012120619410008.html
Published: December 6, 2012.
Japanese text by SciecenNewsline team
based on release from Cell Press
ヨーロッパでは多くの人口を抱えるジプシー「ヨーロッパ・ロマ人」(European Romani)の放浪生活は、
1500年前のインド北西部で始まったことが学術専門誌「Current Biology」を通じて発表された論文により明らかとなった。
今回、発表された論文は、ジプシーの起源に関わる最初の遺伝子調査に基づく研究成果となるだろう。
ジプシーは1100万人の人口を抱え、ヨーロッパの少数派民族グループとしては最大の規模を持つ。
ジプシーの人口は、ギリシャ、ポルトガル、ベルギーといった国の総人口とほとんど変らない。
「私たちは、ヨーロッパの人口において重要な部分を占めているジプシーの人口史を解明することに興味があったのですが、
これまでヨーロッパ各国においてジプシーは無視された存在だったため、彼らに対する科学調査もほとんど行われてきたことはなかったのです」
とスペインのInstitut de Biologia Evolutiva at Universitat Pompeu FabraのDavid Comasは述べる。
ジプシーの起源に関わる歴史書は存在していない。失われたジプシーの歴史を埋めるため、
ComasととErasmus UniversityのManfred Kayserと欧州の国際研究チームは、
ヨーロッパ中から採集された13のジプシー集団の遺伝子情報を使うことにより、ヨーロッパ・ジプシーの起源はインドにあることを確認した。
この結果は、言語学に基づく調査によって得られた従来の学説とも一致するものとなった。
http://jp.sciencenewsline.com/articles/2012120619410008.html
Published: December 6, 2012.
Japanese text by SciecenNewsline team
based on release from Cell Press
ヨーロッパでは多くの人口を抱えるジプシー「ヨーロッパ・ロマ人」(European Romani)の放浪生活は、
1500年前のインド北西部で始まったことが学術専門誌「Current Biology」を通じて発表された論文により明らかとなった。
今回、発表された論文は、ジプシーの起源に関わる最初の遺伝子調査に基づく研究成果となるだろう。
ジプシーは1100万人の人口を抱え、ヨーロッパの少数派民族グループとしては最大の規模を持つ。
ジプシーの人口は、ギリシャ、ポルトガル、ベルギーといった国の総人口とほとんど変らない。
「私たちは、ヨーロッパの人口において重要な部分を占めているジプシーの人口史を解明することに興味があったのですが、
これまでヨーロッパ各国においてジプシーは無視された存在だったため、彼らに対する科学調査もほとんど行われてきたことはなかったのです」
とスペインのInstitut de Biologia Evolutiva at Universitat Pompeu FabraのDavid Comasは述べる。
ジプシーの起源に関わる歴史書は存在していない。失われたジプシーの歴史を埋めるため、
ComasととErasmus UniversityのManfred Kayserと欧州の国際研究チームは、
ヨーロッパ中から採集された13のジプシー集団の遺伝子情報を使うことにより、ヨーロッパ・ジプシーの起源はインドにあることを確認した。
この結果は、言語学に基づく調査によって得られた従来の学説とも一致するものとなった。
>>988
http://en.wikipedia.org/wiki/History_of_the_Romani_people#Genetic_evidence
Genetic evidence[edit]
Researchers doing DNA analysis discovered that Romani populations carried large frequencies of particular Y-chr and mtDNA that otherwise exist only in populations from South Asia.
47.3% of Romani men carry Y-chr of haplogroup H-M82 which is rare outside the South Asia.[22]
mtDNA haplogroup M, most common in Indian subjects and rare outside Southern Asia, accounts for nearly 30% of Romani people.[22]
A more detailed study of Polish Roma shows this to be of the M5 lineage, which is specific to India.[23]
Moreover, a form of the inherited disorder congenital myasthenia is found in Romani subjects.
This form of the disorder, caused by the 1267delG mutation, is otherwise known only in subjects of Indian ancestry.
This is considered to be the best evidence of the Indian ancestry of the Romanies.[24]
A study from 2001 by Gresham et al. suggests "a limited number of related founders, compatible with a small group of migrants splitting from a distinct caste or tribal group".[27]
Also the study pointed out that "genetic drift and different levels and sources of admixture, appear to have played a role in the subsequent differentiation of populations".[27]
The same study found that "a single lineage ... found across Romani populations, accounts for almost one-third of Romani males.
A 2004 study by Morar et al. concluded that the Romanies are "a founder population of common origins that has subsequently split into multiple socially divergent and geographically dispersed Gypsy groups".[24]
The same study revealed that this population "was founded approximately 32?40 generations ago,
with secondary and tertiary founder events occurring approximately 16?25 generations ago".[24]
http://en.wikipedia.org/wiki/History_of_the_Romani_people#Genetic_evidence
Genetic evidence[edit]
Researchers doing DNA analysis discovered that Romani populations carried large frequencies of particular Y-chr and mtDNA that otherwise exist only in populations from South Asia.
47.3% of Romani men carry Y-chr of haplogroup H-M82 which is rare outside the South Asia.[22]
mtDNA haplogroup M, most common in Indian subjects and rare outside Southern Asia, accounts for nearly 30% of Romani people.[22]
A more detailed study of Polish Roma shows this to be of the M5 lineage, which is specific to India.[23]
Moreover, a form of the inherited disorder congenital myasthenia is found in Romani subjects.
This form of the disorder, caused by the 1267delG mutation, is otherwise known only in subjects of Indian ancestry.
This is considered to be the best evidence of the Indian ancestry of the Romanies.[24]
A study from 2001 by Gresham et al. suggests "a limited number of related founders, compatible with a small group of migrants splitting from a distinct caste or tribal group".[27]
Also the study pointed out that "genetic drift and different levels and sources of admixture, appear to have played a role in the subsequent differentiation of populations".[27]
The same study found that "a single lineage ... found across Romani populations, accounts for almost one-third of Romani males.
A 2004 study by Morar et al. concluded that the Romanies are "a founder population of common origins that has subsequently split into multiple socially divergent and geographically dispersed Gypsy groups".[24]
The same study revealed that this population "was founded approximately 32?40 generations ago,
with secondary and tertiary founder events occurring approximately 16?25 generations ago".[24]
990 :世界@名無史さん:2014/02/15(土) 16:06:02.56 0
ところで次スレは?
>>990
おいらはどっかの過疎スレに移るけどね
もともとアーリア人のスレ無くなったから移住しただけだし。
あと、>>979検証中に色々と間違いを発見したんで、
みんなに謝っておきたい。
もう残りのスレ数では修正は不可能だろうな。
ゴメンなさい
おいらはどっかの過疎スレに移るけどね
もともとアーリア人のスレ無くなったから移住しただけだし。
あと、>>979検証中に色々と間違いを発見したんで、
みんなに謝っておきたい。
もう残りのスレ数では修正は不可能だろうな。
ゴメンなさい
お詫びのついでに
最後までお付き合い頂いた方には
これは必見というものをご紹介
>>548
http://www.sciencemag.org/content/342/6155/257/suppl/DC1
http://www.sciencemag.org/content/suppl/2013/10/10/342.6155.257.DC1/1241844s1.mov
まぁ、ちゃんと調べてる人はもう知ってるだろうけどね
ヨーロッパの話は、ほっといても誰かが説明してくれるだろう
と思ったまま3年ぐらいは経っちゃったからなぁ
最後までお付き合い頂いた方には
これは必見というものをご紹介
>>548
http://www.sciencemag.org/content/342/6155/257/suppl/DC1
http://www.sciencemag.org/content/suppl/2013/10/10/342.6155.257.DC1/1241844s1.mov
まぁ、ちゃんと調べてる人はもう知ってるだろうけどね
ヨーロッパの話は、ほっといても誰かが説明してくれるだろう
と思ったまま3年ぐらいは経っちゃったからなぁ
こっちは流石にみんな見てるよね?
>>656のリンク先にあるやつ
原論文
Iosif Lazaridis, Nick Patterson, Alissa Mittnik, et al.
Ancient human genomes suggest three ancestral populations for present-day Europeans
bioRxiv posted online December 23, 2013
doi: 10.1101/001552
http://biorxiv.org/content/early/2013/12/23/001552
後、落ちるまでなら質問やリクエストを受付けるよ。
まぁ、英語は読めないから訳してはあげられないけどねww
>>656のリンク先にあるやつ
原論文
Iosif Lazaridis, Nick Patterson, Alissa Mittnik, et al.
Ancient human genomes suggest three ancestral populations for present-day Europeans
bioRxiv posted online December 23, 2013
doi: 10.1101/001552
http://biorxiv.org/content/early/2013/12/23/001552
後、落ちるまでなら質問やリクエストを受付けるよ。
まぁ、英語は読めないから訳してはあげられないけどねww
>>979
http://www.nature.com/nature/journal/v505/n7481/fig_tab/nature12736_ft.html
http://www.nature.com/nature/journal/v505/n7481/images/nature12736-f1.jpg
a, Geographical locations of Mal’ta and Afontova Gora-2 in south-central Siberia. For reference,
Palaeolithic sites with individuals belonging to mtDNA haplogroup U are shown (red and black triangles):
1, Oberkassel;
2, Hohle Fels;
3, Dolni Vestonice;
4, Kostenki-14.
A Palaeolithic site with an individual belonging to mtDNA haplogroup B is represented by the square:
5, Tianyuan Cave. Notable Palaeolithic sites with Venus figurines are marked by brown circles:
6, Laussel; 7, Lespugue; 8, Grimaldi;
9, Willendorf; 10, Gargarino.
Other notable Palaeolithic sites are shown by grey circles:
11, Sungir; 12, Yana RHS.
b, PCA (PC1 versus PC2) of MA-1 and worldwide human populations for which genomic tracts from recent European admixture in American and Siberian populations have been excluded19.
c, Heat map of the statistic f3(Yoruba; MA-1, X) where X is one of 147 worldwide non-African populations (standard errors shown in Supplementary Fig. 21).
The graded heat key represents the magnitude of the computed f3 statistics.
http://www.nature.com/nature/journal/v505/n7481/images/nature12736-f2.jpg
http://www.nature.com/nature/journal/v505/n7481/fig_tab/nature12736_ft.html
http://www.nature.com/nature/journal/v505/n7481/images/nature12736-f1.jpg
{kind=link}
a, Geographical locations of Mal’ta and Afontova Gora-2 in south-central Siberia. For reference,
Palaeolithic sites with individuals belonging to mtDNA haplogroup U are shown (red and black triangles):
1, Oberkassel;
2, Hohle Fels;
3, Dolni Vestonice;
4, Kostenki-14.
A Palaeolithic site with an individual belonging to mtDNA haplogroup B is represented by the square:
5, Tianyuan Cave. Notable Palaeolithic sites with Venus figurines are marked by brown circles:
6, Laussel; 7, Lespugue; 8, Grimaldi;
9, Willendorf; 10, Gargarino.
Other notable Palaeolithic sites are shown by grey circles:
11, Sungir; 12, Yana RHS.
b, PCA (PC1 versus PC2) of MA-1 and worldwide human populations for which genomic tracts from recent European admixture in American and Siberian populations have been excluded19.
c, Heat map of the statistic f3(Yoruba; MA-1, X) where X is one of 147 worldwide non-African populations (standard errors shown in Supplementary Fig. 21).
The graded heat key represents the magnitude of the computed f3 statistics.
http://www.nature.com/nature/journal/v505/n7481/images/nature12736-f2.jpg
{kind=link}
995 :世界@名無史さん:
示された論文を読む限りでは、
欧州人の遺伝子的な歴史を整理するにはまだ時間がかかるということだね。
現代欧州人の起源が3種類の「集団」から成るというのは定説化しつつあるが、
それに遺伝子・人種形態・語族・分化・侵入経路をどう組み合わせていくかというのは、
大変労力がいるし、まだまだサンプルが必要だね。
ただ、北部欧州人とNATIVE AMERICANとの類縁性については、
かえって頭を混乱させる学説だね。
次スレでもよろしく。
欧州人の遺伝子的な歴史を整理するにはまだ時間がかかるということだね。
現代欧州人の起源が3種類の「集団」から成るというのは定説化しつつあるが、
それに遺伝子・人種形態・語族・分化・侵入経路をどう組み合わせていくかというのは、
大変労力がいるし、まだまだサンプルが必要だね。
ただ、北部欧州人とNATIVE AMERICANとの類縁性については、
かえって頭を混乱させる学説だね。
次スレでもよろしく。